

Coordination of mitochondrial bioenergetics with G1 phase cell cycle progression
- PMID: 18583942
- PMCID: PMC3399174
- DOI: 10.4161/cc.7.12.6067
Coordination of mitochondrial bioenergetics with G1 phase cell cycle progression
Abstract
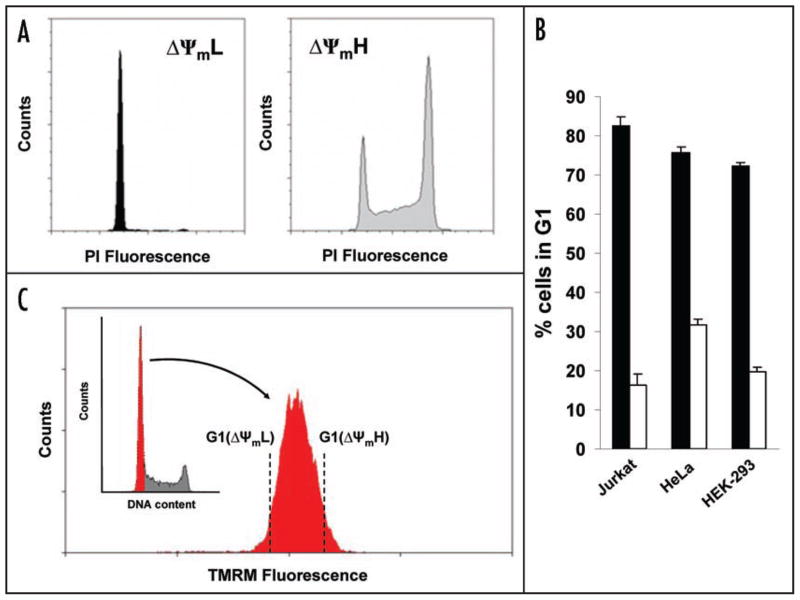
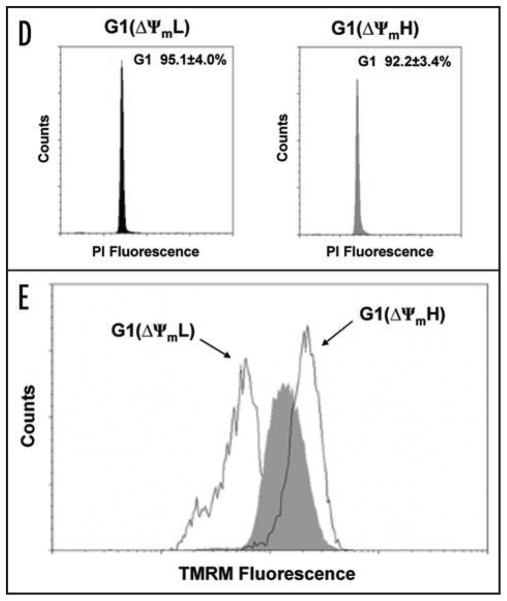
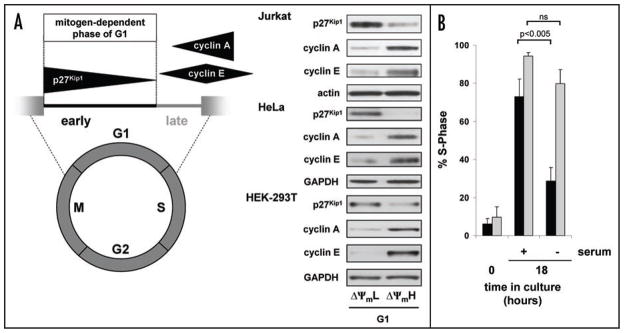
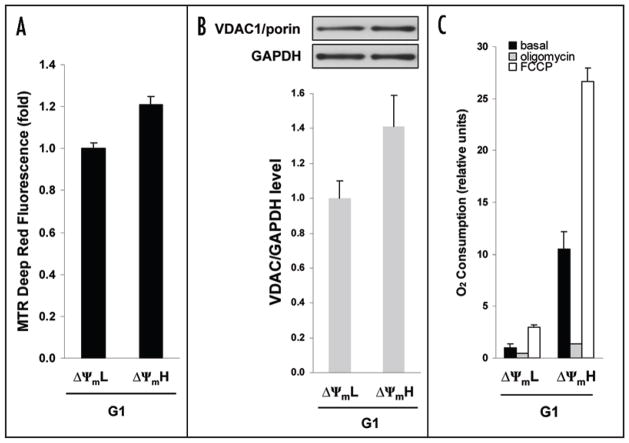
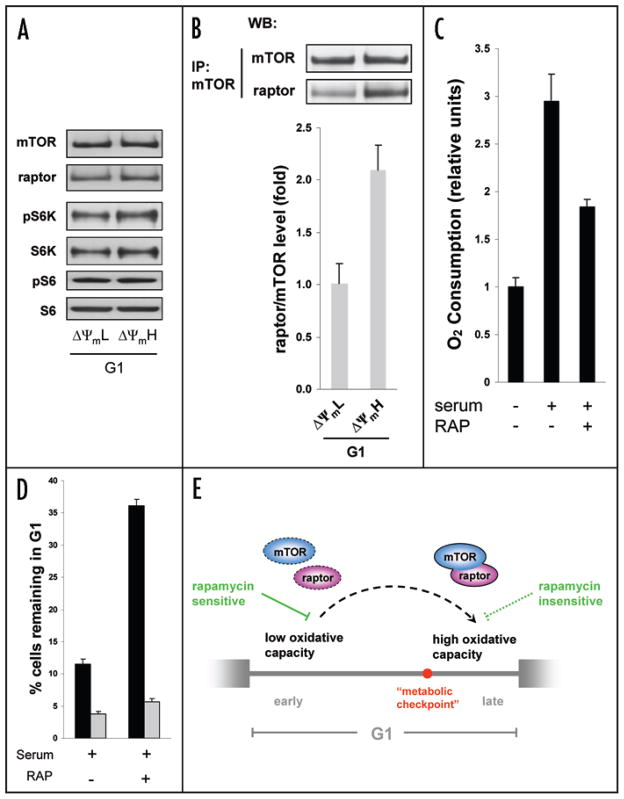
Relatively little is known regarding how energetic demand during cell proliferation is sensed or coordinated with mitochondrial metabolism. Here we demonstrate that cell cycle progression through G(1) is associated with a significant increase in mitochondrial membrane potential (DeltaPsi(m)) and respiration. We used this change in metabolic rate to isolate cells in G(1) with low and high levels of mitochondrial membrane potential (DeltaPsi(m)L and DeltaPsi(m)H). Biochemical and functional studies demonstrate that DeltaPsi(m)L and DeltaPsi(m)H cells display the distinct characteristics of early and late G(1) phase, respectively. We further demonstrate that the metabolic rate in G(1) reflects levels of the mTOR-raptor complex as well as susceptibility to rapamycin-induced cell cycle delay. In conclusion, our data suggests a coupling of mitochondrial bioenergetics and G(1) progression and points to the mTOR signaling pathway as a potential molecular coordinator of these two processes.
Figures
References
-
- Mandal S, Guptan P, Owusu-Ansah E, Banerjee U. Mitochondrial regulation of cell cycle progression during development as revealed by the tenured mutation in Drosophila. Dev Cell. 2005;9:843–54. - PubMed
-
- Jones RG, Plas DR, Kubek S, Buzzai M, Mu J, Xu Y, Birnbaum MJ, Thompson CB. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol Cell. 2005;18:283–93. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous