

(-)-Epigallocatechin gallate causes internalization of the epidermal growth factor receptor in human colon cancer cells
- PMID: 18586691
- PMCID: PMC2556969
- DOI: 10.1093/carcin/bgn128
(-)-Epigallocatechin gallate causes internalization of the epidermal growth factor receptor in human colon cancer cells
Abstract
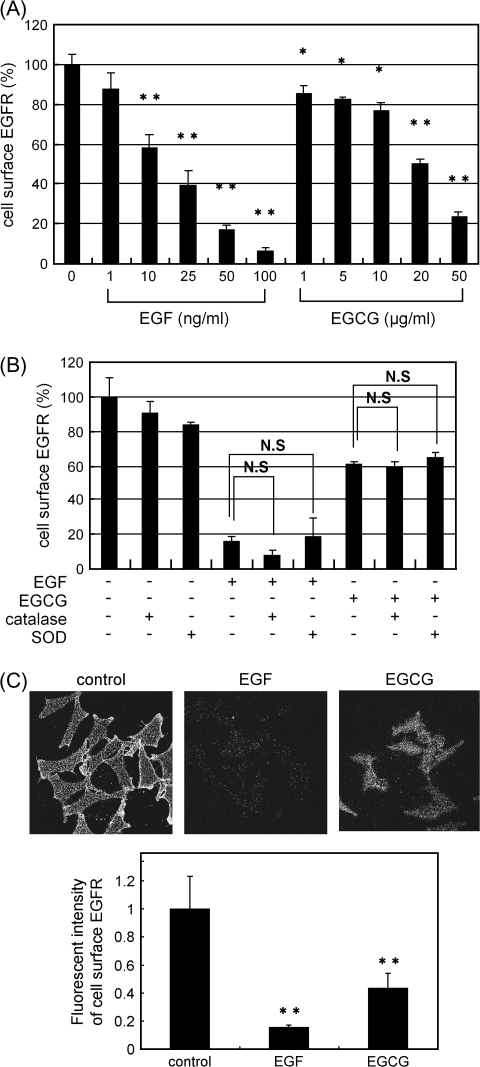
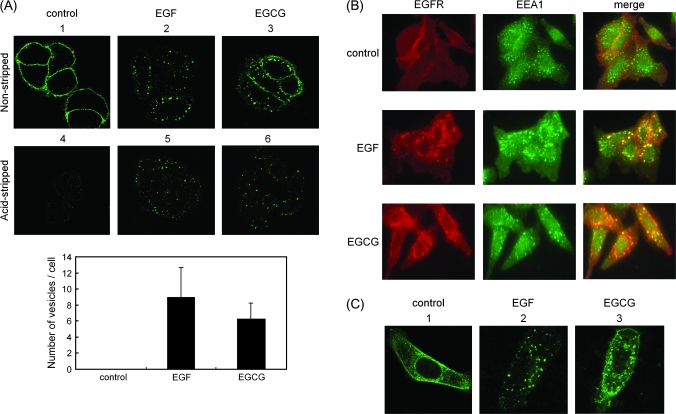
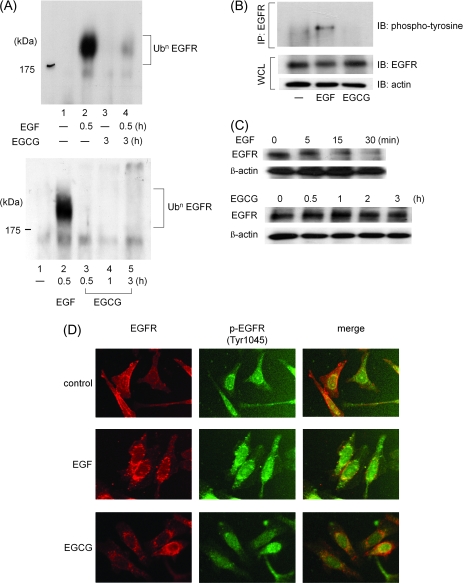
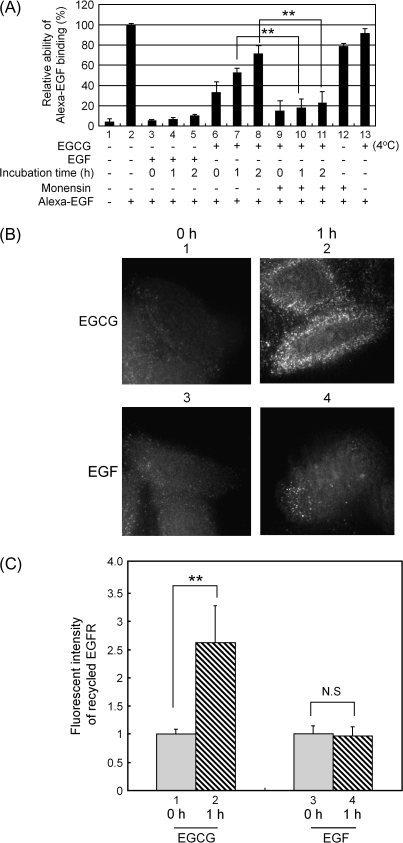
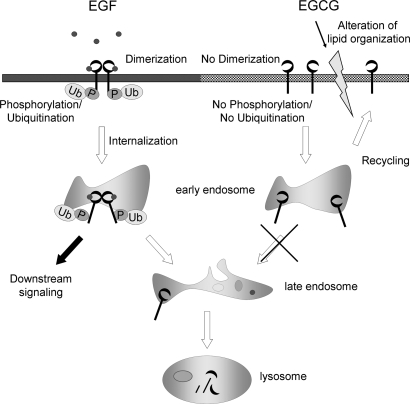
We recently found that the inhibitory effect of (-)-epigallocatechin gallate (EGCG) on epidermal growth factor (EGF) binding to the epidermal growth factor receptor (EGFR) is associated with alterations in lipid organization in the plasma membrane of colon cancer cells. Since changes in lipid organizations are thought to play a role in the trafficking of several membrane proteins, in this study we examined the effects of EGCG on cellular localization of the EGFR in SW480 cells. Treatment of the cells for 30 min with as little as 1 microg/ml of EGCG caused a decrease in cell surface-associated EGFRs and this was associated with internalization of EGFRs into endosomal vesicles. Similar effects were seen with a green fluorescent protein (GFP)-EGFR fusion protein. As expected, the EGFR protein was phosphorylated at tyrosine residues, ubiquitinated and partially degraded when the cells were treated with EGF, but treatment with EGCG caused none of these effects. The loss of EGFRs from the cell surface induced by treating the cells with EGF for 30 min persisted for at least 2 h. However, the loss of EGFRs from the cell surface induced by temporary exposure to EGCG was partially restored within 1-2 h. These studies provide the first evidence that EGCG can induce internalization of EGFRs into endosomes, which can recycle back to the cell surface. This sequestrating of inactivated EGFRs into endosomes may explain, at least in part, the ability of EGCG to inhibit activation of the EGFR and thereby exert anticancer effects.
Figures
References
-
- Masuda M, et al. Effects of epigallocatechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin. Cancer Res. 2001;7:4220–4229. - PubMed
-
- Pianetti S, et al. Green tea polyphenol epigallocatechin-3 gallate inhibits Her-2/neu signaling, proliferation, and transformed phenotype of breast cancer cells. Cancer Res. 2002;62:652–655. - PubMed
-
- Paschka AG, et al. Induction of apoptosis in prostate cancer cell lines by the green tea component, (-)-epigallocatechin-3-gallate. Cancer Lett. 1998;130:1–7. - PubMed
-
- Shimizu M, et al. (-)-Epigallocatechin gallate and polyphenon E inhibit growth and activation of the epidermal growth factor receptor and human epidermal growth factor receptor-2 signaling pathways in human colon cancer cells. Clin. Cancer Res. 2005;11:2735–2746. - PubMed
-
- Khan N, et al. Targeting multiple signaling pathways by green tea polyphenol (-)-epigallocatechin-3-gallate. Cancer Res. 2006;66:2500–2505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
