

Different requirements of the kinase and UHM domains of KIS for its nuclear localization and binding to splicing factors
- PMID: 18588901
- PMCID: PMC2632974
- DOI: 10.1016/j.jmb.2008.06.026
Different requirements of the kinase and UHM domains of KIS for its nuclear localization and binding to splicing factors
Abstract
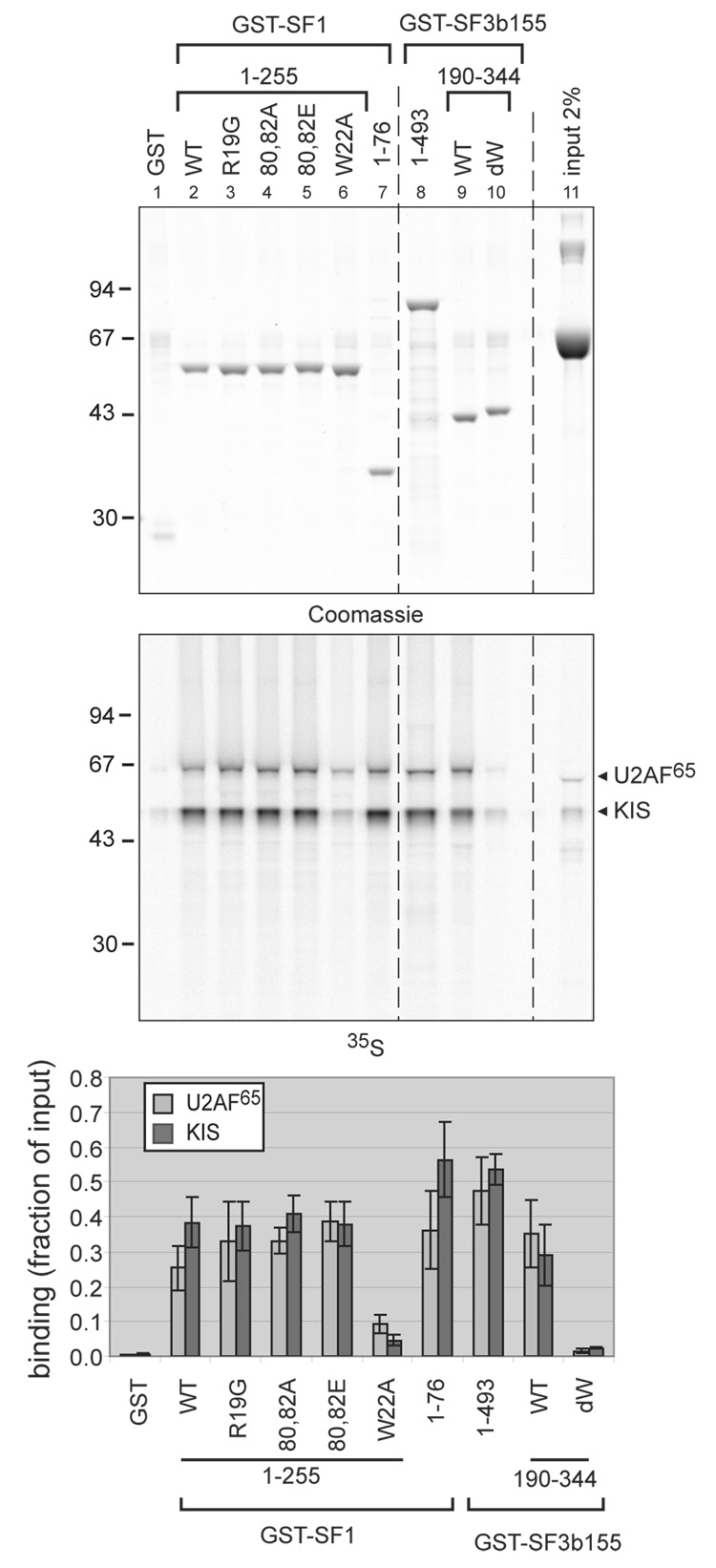
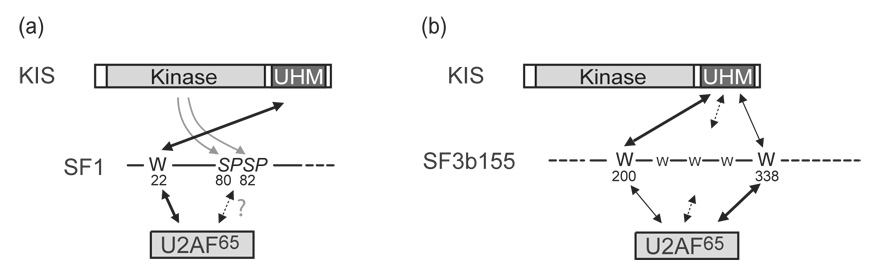
The protein kinase KIS is made by the juxtaposition of a unique kinase domain and a C-terminal domain with a U2AF homology motif (UHM), a sequence motif for protein interaction initially identified in the heterodimeric pre-mRNA splicing factor U2AF. This domain of KIS is closely related to the C-terminal UHM domain of the U2AF large subunit, U2AF(65). KIS phosphorylates the splicing factor SF1, which in turn enhances SF1 binding to U2AF(65) and the 3' splice site, an event known to take place at an early step of spliceosome assembly. Here, the analysis of the subcellular localization of mutated forms of KIS indicates that the kinase domain of KIS is the necessary domain for its nuclear localization. As in the case of U2AF(65), the UHM-containing C-terminal domain of KIS is required for binding to the splicing factors SF1 and SF3b155. The efficiency of KIS binding to SF1 and SF3b155 is similar to that of U2AF(65) in pull-down assays. These results further support the functional link of KIS with splicing factors. Interestingly, when compared to other UHM-containing proteins, KIS presents a different specificity for the UHM docking sites that are present in the N-terminal region of SF3b155, thus providing a new insight into the variety of interactions mediated by UHM domains.
Figures






References
-
- Hunter T. Signaling--2000 and beyond. Cell. 2000;100:113–127. - PubMed
-
- Maris C, Dominguez C, Allain FH. The RNA recognition motif, a plastic RNA-binding platform to regulate post-transcriptional gene expression. Febs J. 2005;272:2118–2131. - PubMed
-
- Maucuer A, Ozon S, Manceau V, Gavet O, Lawler S, Curmi P, Sobel A. KIS is a protein kinase with an RNA recognition motif. J Biol Chem. 1997;272:23151–23156. - PubMed
-
- Bieche I, Manceau V, Curmi PA, Laurendeau I, Lachkar S, Leroy K, Vidaud D, Sobel A, Maucuer A. Quantitative RT-PCR reveals a ubiquitous but preferentially neural expression of the KIS gene in rat and human. Brain Res Mol Brain Res. 2003;114:55–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
