

The SOCS box domain of SOCS3: structure and interaction with the elonginBC-cullin5 ubiquitin ligase
- PMID: 18590740
- PMCID: PMC3652878
- DOI: 10.1016/j.jmb.2008.06.038
The SOCS box domain of SOCS3: structure and interaction with the elonginBC-cullin5 ubiquitin ligase
Abstract
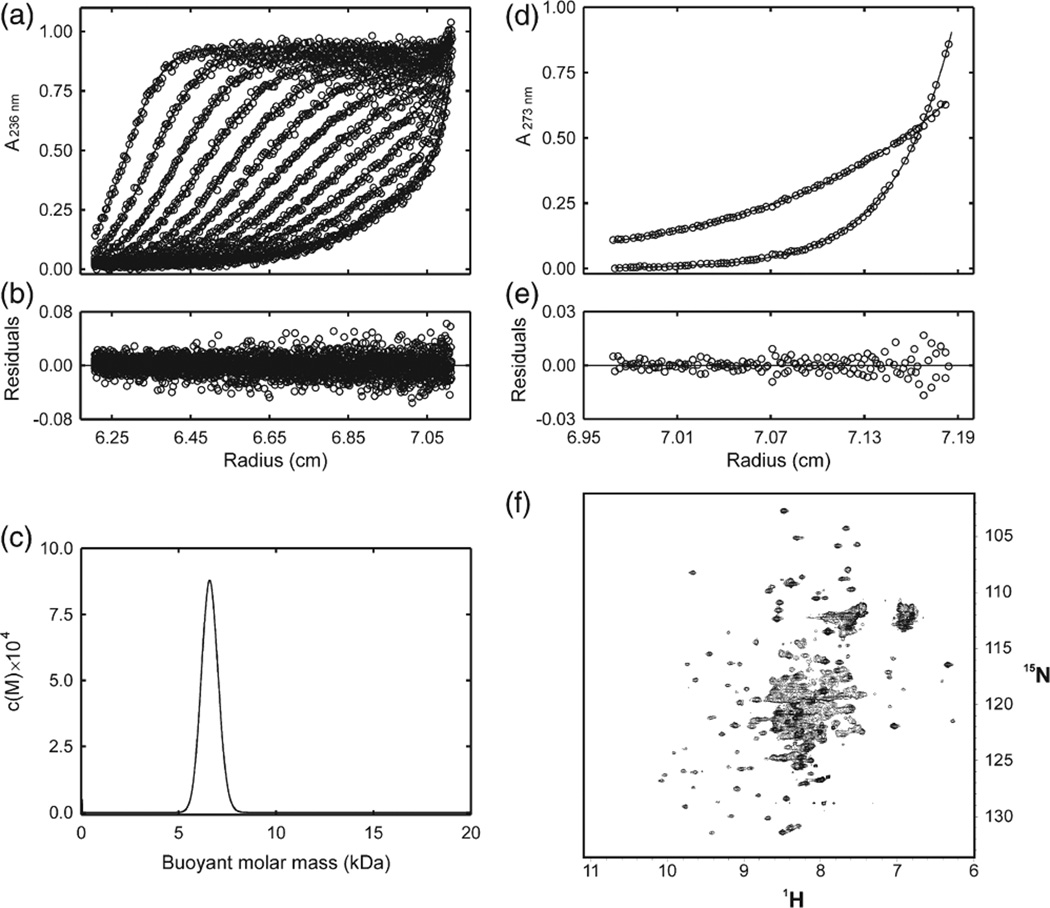
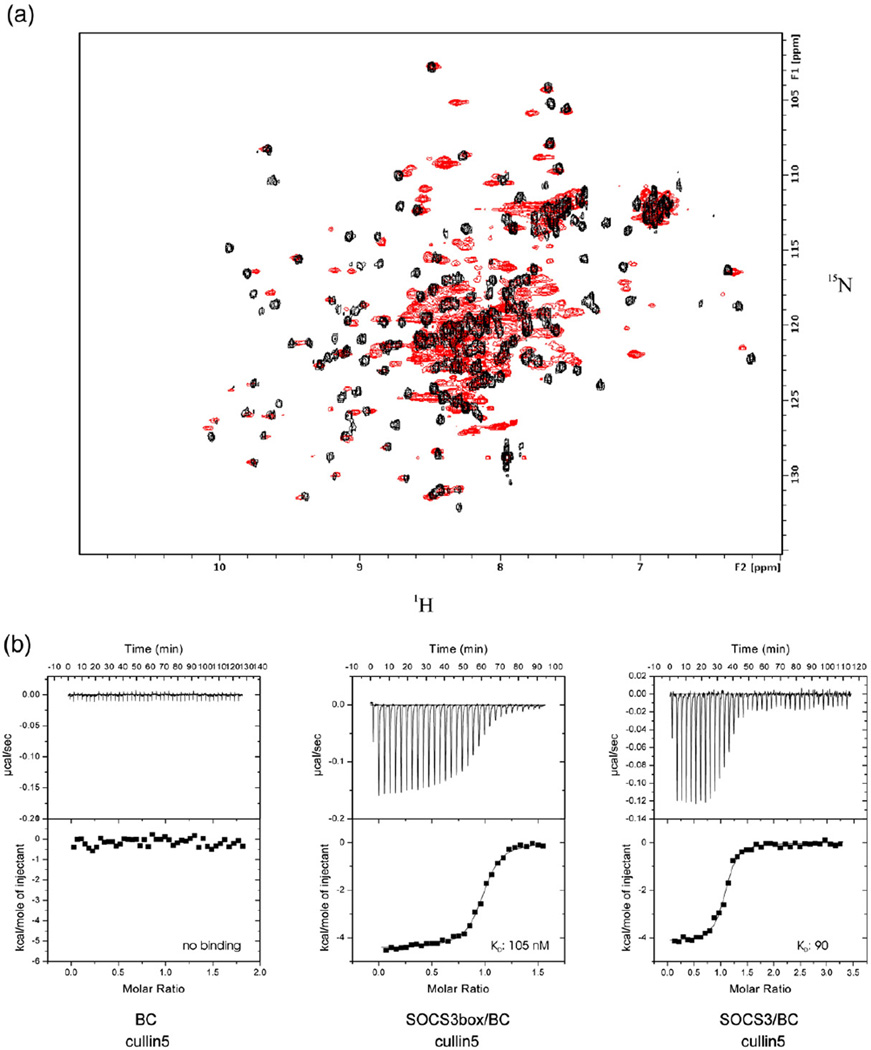
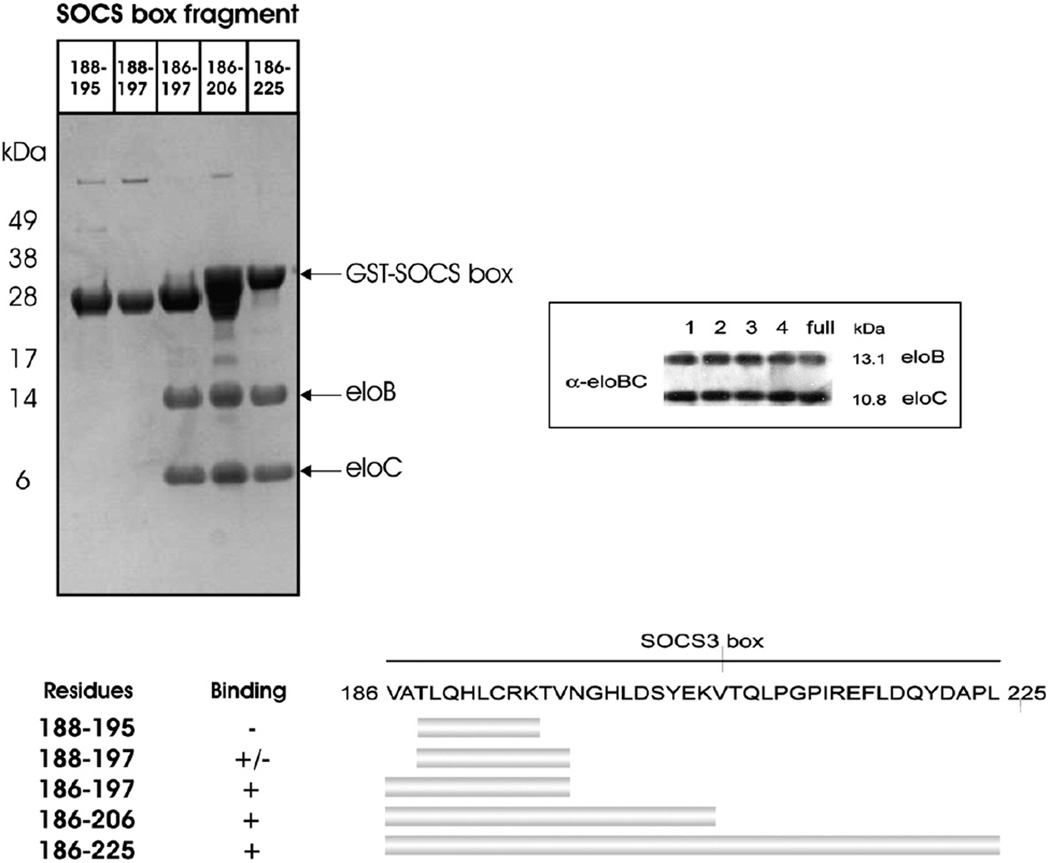
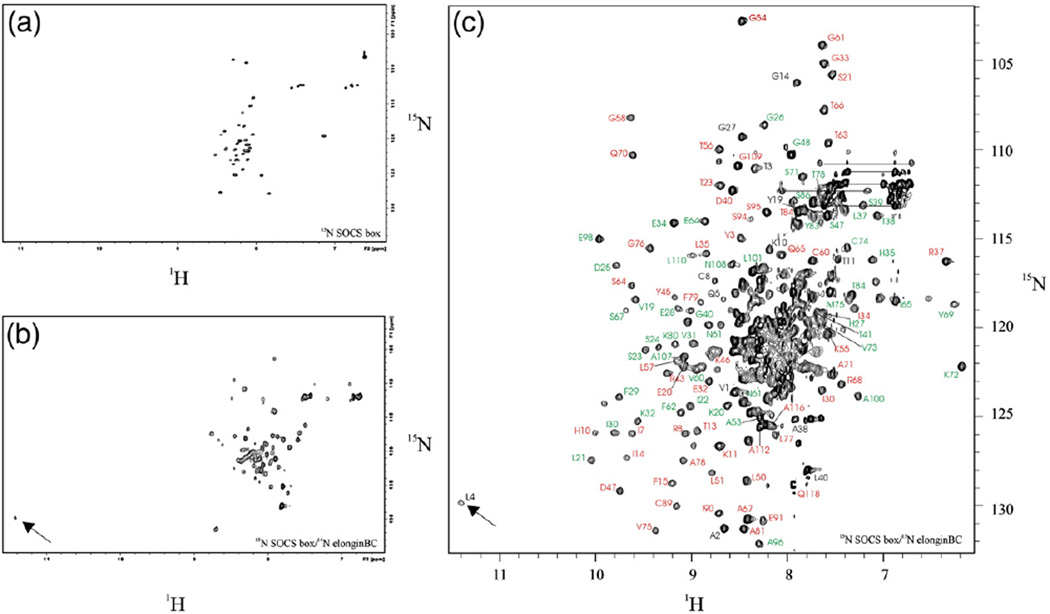
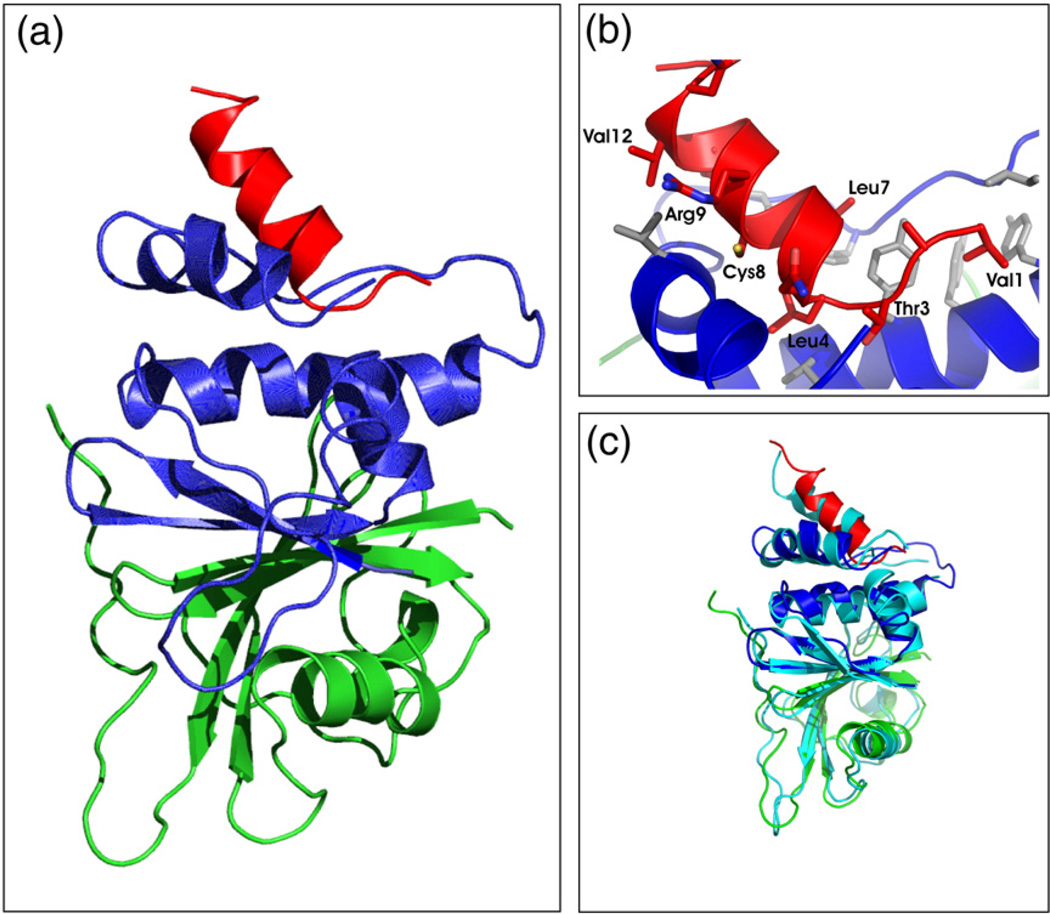
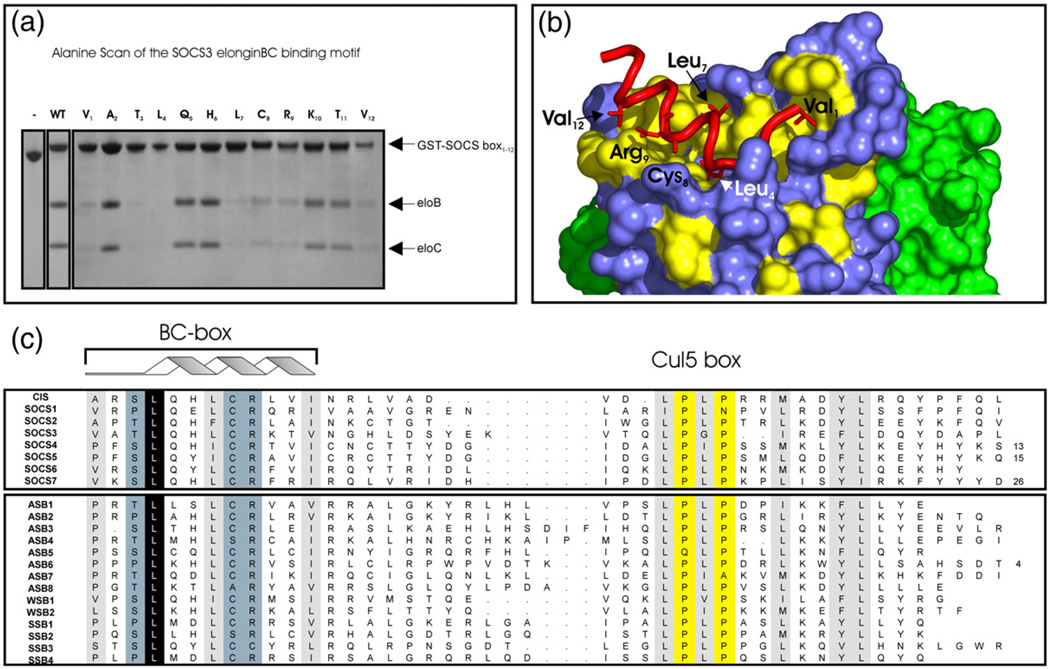
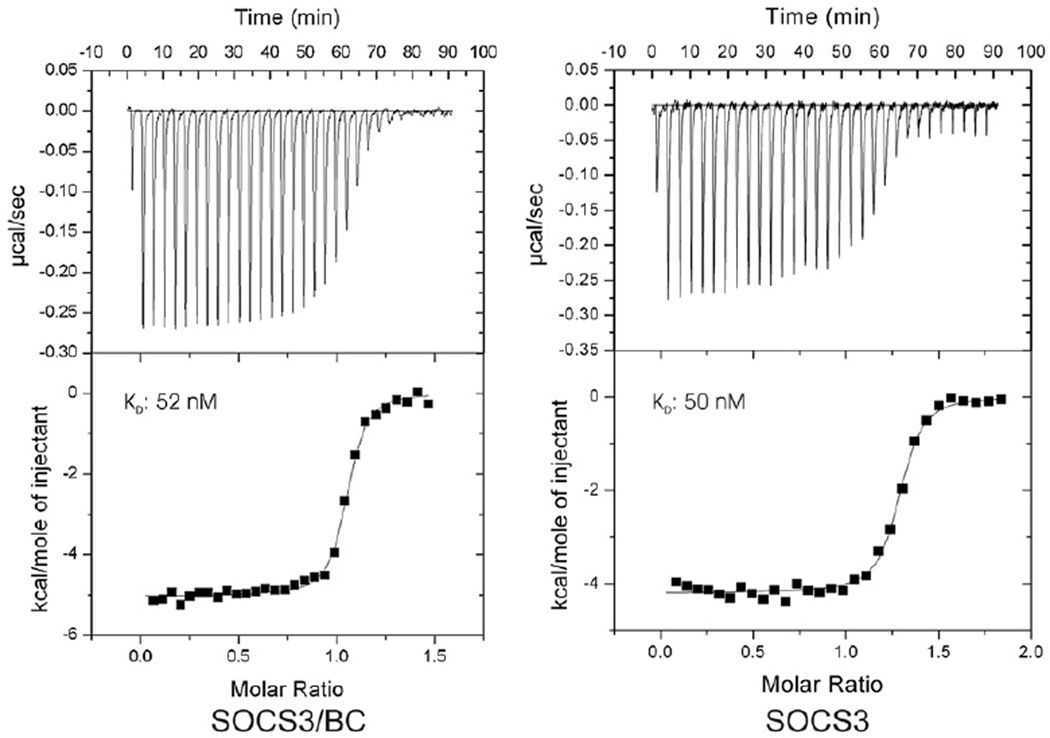
Suppressor of cytokine signalling 3 (SOCS3) is responsible for regulating the cellular response to a variety of cytokines, including interleukin 6 and leukaemia inhibitory factor. Identification of the SOCS box domain led to the hypothesis that SOCS3 can associate with functional E3 ubiquitin ligases and thereby induce the degradation of bound signalling proteins. This model relies upon an interaction between the SOCS box, elonginBC and a cullin protein that forms the E3 ligase scaffold. We have investigated this interaction in vitro using purified components and show that SOCS3 binds to elonginBC and cullin5 with high affinity. The SOCS3-elonginBC interaction was further characterised by determining the solution structure of the SOCS box-elonginBC ternary complex and by deletion and alanine scanning mutagenesis of the SOCS box. These studies revealed that conformational flexibility is a key feature of the SOCS-elonginBC interaction. In particular, the SOCS box is disordered in isolation and only becomes structured upon elonginBC association. The interaction depends upon the first 12 residues of the SOCS box domain and particularly on a deeply buried, conserved leucine. The SOCS box, when bound to elonginBC, binds tightly to cullin5 with 100 nM affinity. Domains upstream of the SOCS box are not required for elonginBC or cullin5 association, indicating that the SOCS box acts as an independent binding domain capable of recruiting elonginBC and cullin5 to promote E3 ligase formation.
Figures
References
-
- Starr R, Willson TA, Viney EM, Murray LJ, Rayner JR, Jenkins BJ, et al. A family of cytokine-inducible inhibitors of signalling. Nature. 1997;387:917–921. - PubMed
-
- Leonard WJ, O’Shea JJ. Jaks and STATs: biological implications. Annu. Rev. Immunol. 1998;16:293–322. - PubMed
-
- Kimura A, Kinjyo I, Matsumura Y, Mori H, Mashima R, Harada M, et al. SOCS3 is a physiological negative regulator for granulopoiesis and granulocyte colony-stimulating factor receptor signaling. J. Biol. Chem. 2004;279:6905–6910. - PubMed
-
- Croker BA, Metcalf D, Robb L, Wei W, Mifsud S, DiRago L, et al. SOCS3 is a critical physiological negative regulator of G-CSF signaling and emergency granulopoiesis. Immunity. 2004;20:153–165. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
