

Inhibition of apoptosome formation by suppression of Hsp90beta phosphorylation in tyrosine kinase-induced leukemias
- PMID: 18591256
- PMCID: PMC2519729
- DOI: 10.1128/MCB.00265-08
Inhibition of apoptosome formation by suppression of Hsp90beta phosphorylation in tyrosine kinase-induced leukemias
Abstract
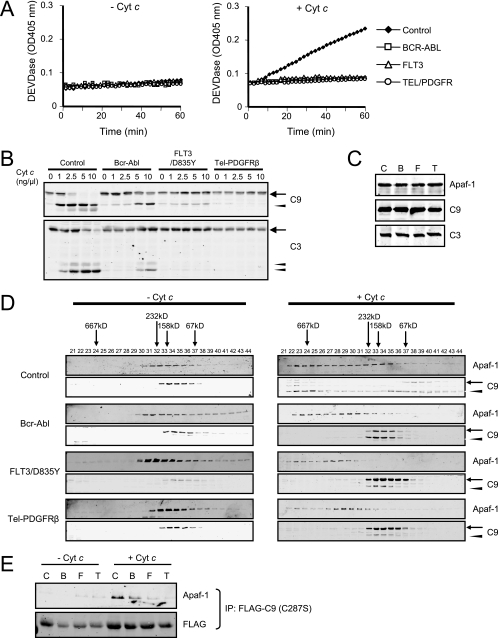
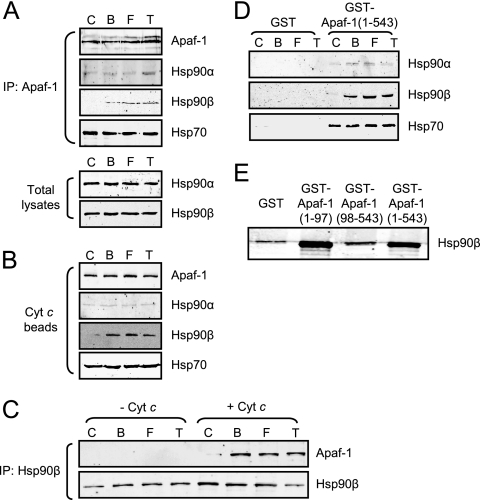
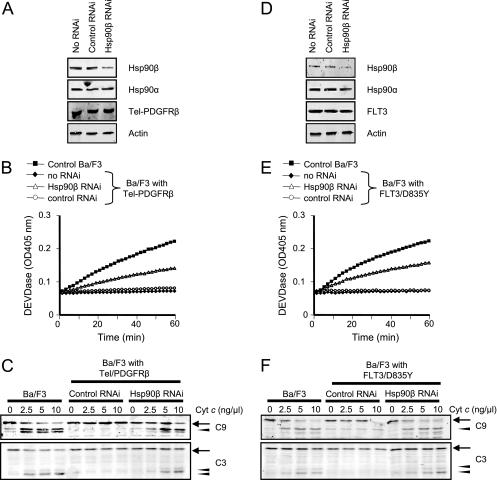
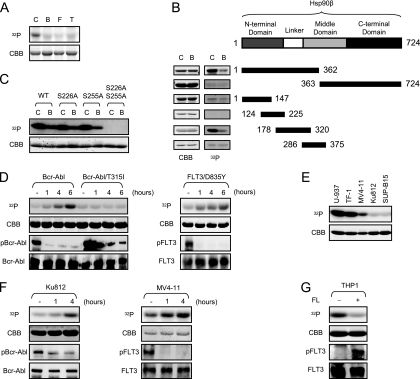
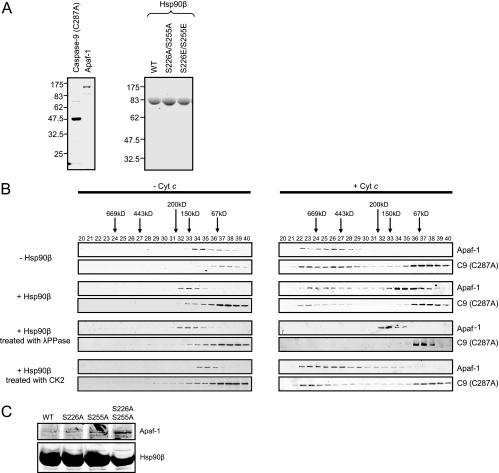
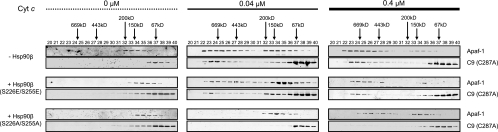
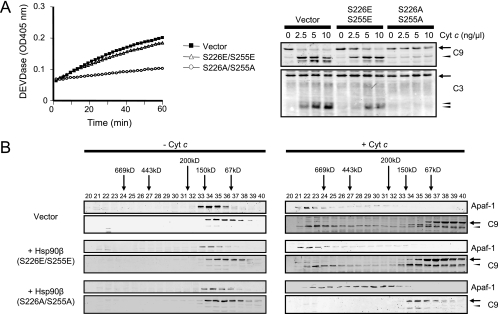
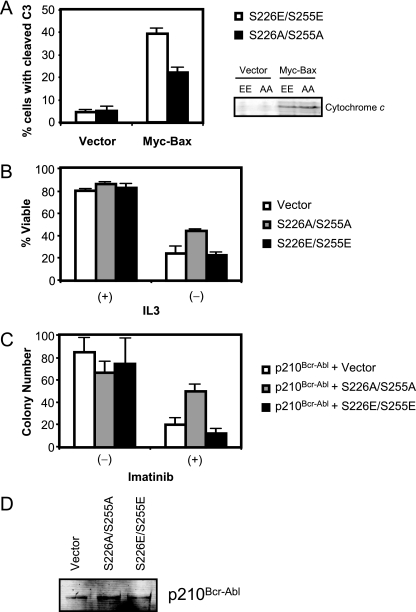
Constitutively active tyrosine kinases promote leukemogenesis by increasing cell proliferation and inhibiting apoptosis. However, mechanisms underlying apoptotic inhibition have not been fully elucidated. In many settings, apoptosis occurs by mitochondrial cytochrome c release, which nucleates the Apaf-1/caspase-9 apoptosome. Here we report that the leukemogenic kinases, Bcr-Abl, FLT3/D835Y, and Tel-PDGFRbeta, all can inhibit apoptosome function. In cells expressing these kinases, the previously reported apoptosome inhibitor, Hsp90beta, bound strongly to Apaf-1, preventing cytochrome c-induced Apaf-1 oligomerization and caspase-9 recruitment. Hsp90beta interacted weakly with the apoptosome in untransformed cells. While Hsp90beta was phosphorylated at Ser 226/Ser 255 in untransformed cells, phosphorylation was absent in leukemic cells. Expression of mutant Hsp90beta (S226A/S255A), which mimics the hypophosphorylated form in leukemic cells, conferred resistance to cytochrome c-induced apoptosome activation in normal cells, reflecting enhanced binding of nonphosphorylatable Hsp90beta to Apaf-1. In Bcr-Abl-positive mouse bone marrow cells, nonphosphorylatable Hsp90beta expression conferred imatinib (Gleevec) resistance. These data provide an explanation for apoptosome inhibition by activated leukemogenic tyrosine kinases and suggest that alterations in Hsp90beta-apoptosome interactions may contribute to chemoresistance in leukemias.
Figures
Similar articles
-
Dihydroartemisinin inhibits the Bcr/Abl oncogene at the mRNA level in chronic myeloid leukemia sensitive or resistant to imatinib.Biomed Pharmacother. 2013 Mar;67(2):157-63. doi: 10.1016/j.biopha.2012.10.017. Epub 2012 Nov 19. Biomed Pharmacother. 2013. PMID: 23201011
-
Disruption of the Bcr-Abl/Hsp90 protein complex: a possible mechanism to inhibit Bcr-Abl-positive human leukemic blasts by novobiocin.Leukemia. 2008 Jul;22(7):1402-9. doi: 10.1038/leu.2008.89. Epub 2008 Apr 17. Leukemia. 2008. Retraction in: Leukemia. 2020 Jun;34(6):1722. doi: 10.1038/s41375-020-0823-z. PMID: 18418407 Retracted.
-
ARG tyrosine kinase activity is inhibited by STI571.Blood. 2001 Apr 15;97(8):2440-8. doi: 10.1182/blood.v97.8.2440. Blood. 2001. PMID: 11290609
-
[The role of the apoptosome in the activation of procaspase-9].Postepy Hig Med Dosw (Online). 2013 Feb 6;67:54-64. doi: 10.5604/17322693.1032333. Postepy Hig Med Dosw (Online). 2013. PMID: 23475483 Review. Polish.
-
A molecular view on signal transduction by the apoptosome.Cell Signal. 2012 Jul;24(7):1420-5. doi: 10.1016/j.cellsig.2012.03.007. Epub 2012 Mar 13. Cell Signal. 2012. PMID: 22446004 Review.
Cited by
-
Triptolide, a HSP90 middle domain inhibitor, induces apoptosis in triple manner.Oncotarget. 2018 Apr 27;9(32):22301-22315. doi: 10.18632/oncotarget.24737. eCollection 2018 Apr 27. Oncotarget. 2018. PMID: 29854279 Free PMC article.
-
FNDC4 acts as an anti-inflammatory factor on macrophages and improves colitis in mice.Nat Commun. 2016 Apr 12;7:11314. doi: 10.1038/ncomms11314. Nat Commun. 2016. PMID: 27066907 Free PMC article.
-
The levels of retinoic acid-inducible gene I are regulated by heat shock protein 90-alpha.J Immunol. 2009 Mar 1;182(5):2717-25. doi: 10.4049/jimmunol.0802933. J Immunol. 2009. PMID: 19234166 Free PMC article.
-
Hsp-90 and the biology of nematodes.BMC Evol Biol. 2009 Oct 22;9:254. doi: 10.1186/1471-2148-9-254. BMC Evol Biol. 2009. PMID: 19849843 Free PMC article.
-
Protective Role of Genetic Variants in HSP90 Genes-Complex in COPD Secondary to Biomass-Burning Smoke Exposure and Non-Severe COPD Forms in Tobacco Smoking Subjects.Curr Issues Mol Biol. 2021 Aug 3;43(2):887-899. doi: 10.3390/cimb43020063. Curr Issues Mol Biol. 2021. PMID: 34449539 Free PMC article.
References
-
- Abu-Duhier, F. M., A. C. Goodeve, G. A. Wilson, R. S. Care, I. R. Peake, and J. T. Reilly. 2001. Identification of novel FLT-3 Asp835 mutations in adult acute myeloid leukaemia. Br. J. Haematol. 113983-988. - PubMed
-
- Amarante-Mendes, G. P., A. J. McGahon, W. K. Nishioka, D. E. Afar, O. N. Witte, and D. R. Green. 1998. Bcl-2-independent Bcr-Abl-mediated resistance to apoptosis: protection is correlated with up regulation of Bcl-xL. Oncogene 161383-1390. - PubMed
-
- An, W. G., T. W. Schulte, and L. M. Neckers. 2000. The heat shock protein 90 antagonist geldanamycin alters chaperone association with p210bcr-abl and v-src proteins before their degradation by the proteasome. Cell Growth Differ. 11355-360. - PubMed
-
- Bhatia, R., M. Holtz, N. Niu, R. Gray, D. S. Snyder, C. L. Sawyers, D. A. Arber, M. L. Slovak, and S. J. Forman. 2003. Persistence of malignant hematopoietic progenitors in chronic myelogenous leukemia patients in complete cytogenetic remission following imatinib mesylate treatment. Blood 1014701-4707. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous