

Clinical, agricultural, and evolutionary biology of myostatin: a comparative review
- PMID: 18591260
- PMCID: PMC2528853
- DOI: 10.1210/er.2008-0003
Clinical, agricultural, and evolutionary biology of myostatin: a comparative review
Abstract
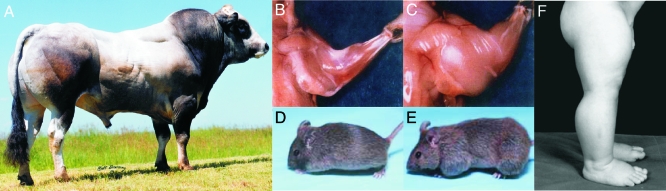
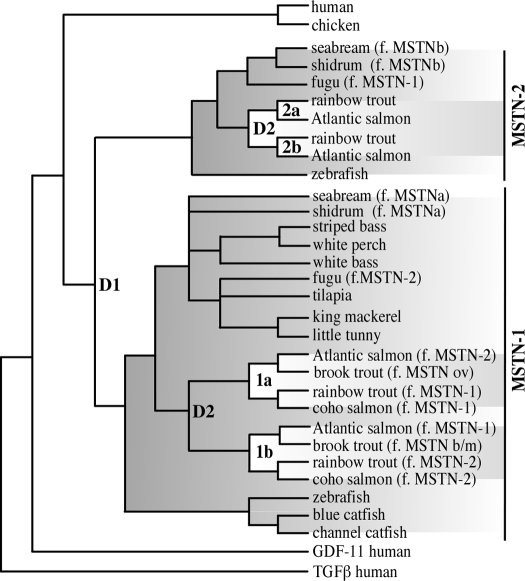
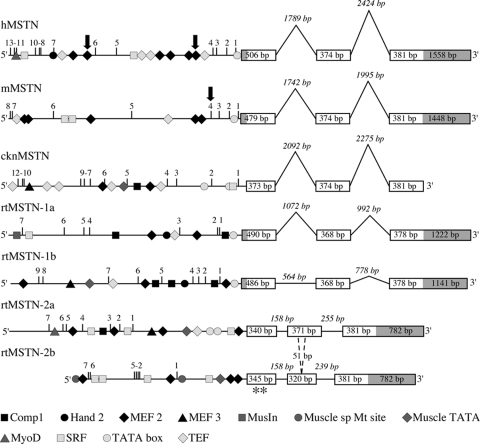
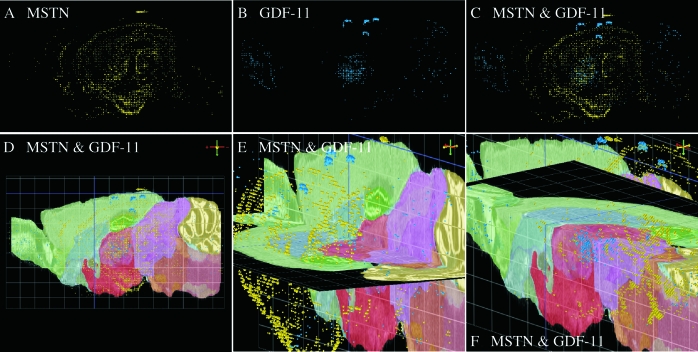
The discovery of myostatin and our introduction to the "Mighty Mouse" over a decade ago spurred both basic and applied research and impacted popular culture as well. The myostatin-null genotype produces "double muscling" in mice and livestock and was recently described in a child. The field's rapid growth is by no means surprising considering the potential benefits of enhancing muscle growth in clinical and agricultural settings. Indeed, several recent studies suggest that blocking myostatin's inhibitory effects could improve the clinical treatment of several muscle growth disorders, whereas comparative studies suggest that these actions are at least partly conserved. Thus, neutralizing myostatin's effects could also have agricultural significance. Extrapolating between studies that use different vertebrate models, particularly fish and mammals, is somewhat confusing because whole genome duplication events have resulted in the production and retention of up to four unique myostatin genes in some fish species. Such comparisons, however, suggest that myostatin's actions may not be limited to skeletal muscle per se, but may additionally influence other tissues including cardiac muscle, adipocytes, and the brain. Thus, therapeutic intervention in the clinic or on the farm must consider the potential of alternative side effects that could impact these or other tissues. In addition, the presence of multiple and actively diversifying myostatin genes in most fish species provides a unique opportunity to study adaptive molecular evolution. It may also provide insight into myostatin's nonmuscle actions as results from these and other comparative studies gain visibility in biomedical fields.
Figures


References
-
- McPherron AC, Lawler AM, Lee SJ 1997 Regulation of skeletal muscle mass in mice by a new TGF-β superfamily member. Nature 387:83–90 - PubMed
-
- Grobet L, Martin LJ, Poncelet D, Pirottin D, Brouwers B, Riquet J, Schoeberlein A, Dunner S, Menissier F, Massabanda J, Fries R, Hanset R, Georges M 1997 A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle [see comments]. Nat Genet 17:71–74 - PubMed
-
- Kambadur R, Sharma M, Smith TP, Bass JJ 1997 Mutations in myostatin (GDF8) in double-muscled Belgian Blue and Piedmontese cattle. Genome Res 7:910–916 - PubMed
-
- Smith TP, Lopez-Corrales NL, Kappes SM, Sonstegard TS 1997 Myostatin maps to the interval containing the bovine mh locus. Mamm Genome 8:742–744 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
