

Cytokine-induced beta-cell death is independent of endoplasmic reticulum stress signaling
- PMID: 18591394
- PMCID: PMC2570400
- DOI: 10.2337/db07-1802
Cytokine-induced beta-cell death is independent of endoplasmic reticulum stress signaling
Abstract
Objective: Cytokines contribute to beta-cell destruction in type 1 diabetes. Endoplasmic reticulum (ER) stress-mediated apoptosis has been proposed as a mechanism for beta-cell death. We tested whether ER stress was necessary for cytokine-induced beta-cell death and also whether ER stress gene activation was present in beta-cells of the NOD mouse model of type 1 diabetes.
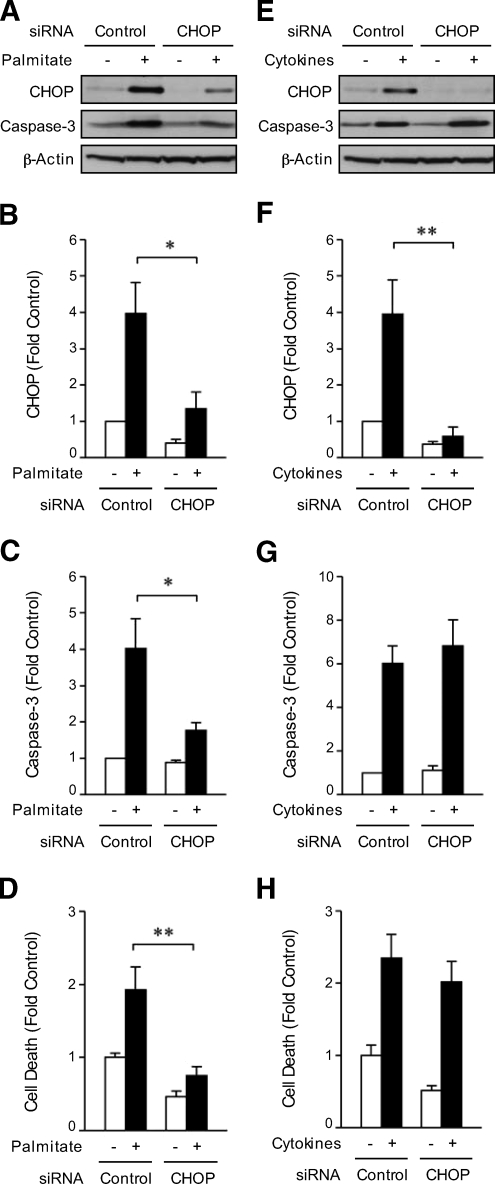
Research design and methods: INS-1 beta-cells or rat islets were treated with the chemical chaperone phenyl butyric acid (PBA) and exposed or not to interleukin (IL)-1beta and gamma-interferon (IFN-gamma). Small interfering RNA (siRNA) was used to silence C/EBP homologous protein (CHOP) expression in INS-1 beta-cells. Additionally, the role of ER stress in lipid-induced cell death was assessed.
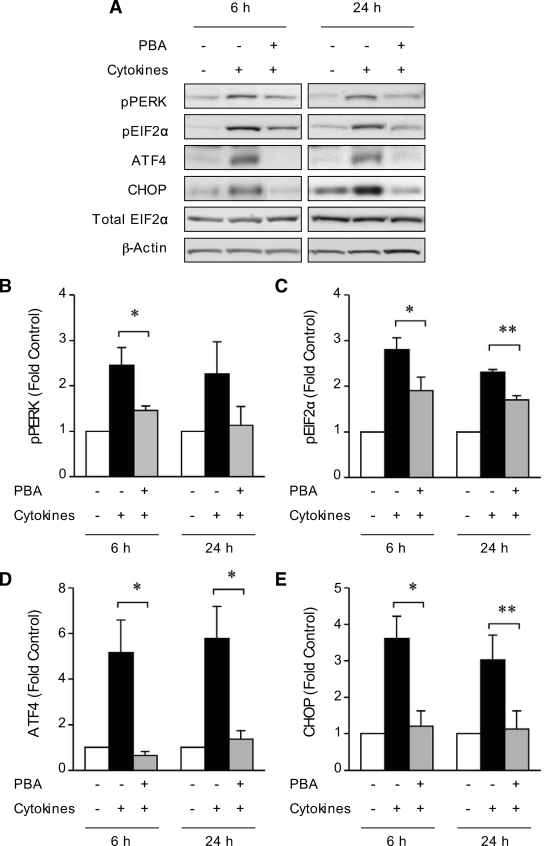
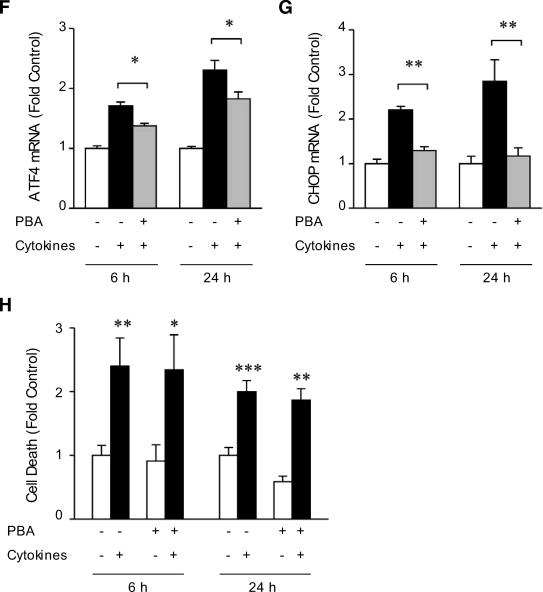
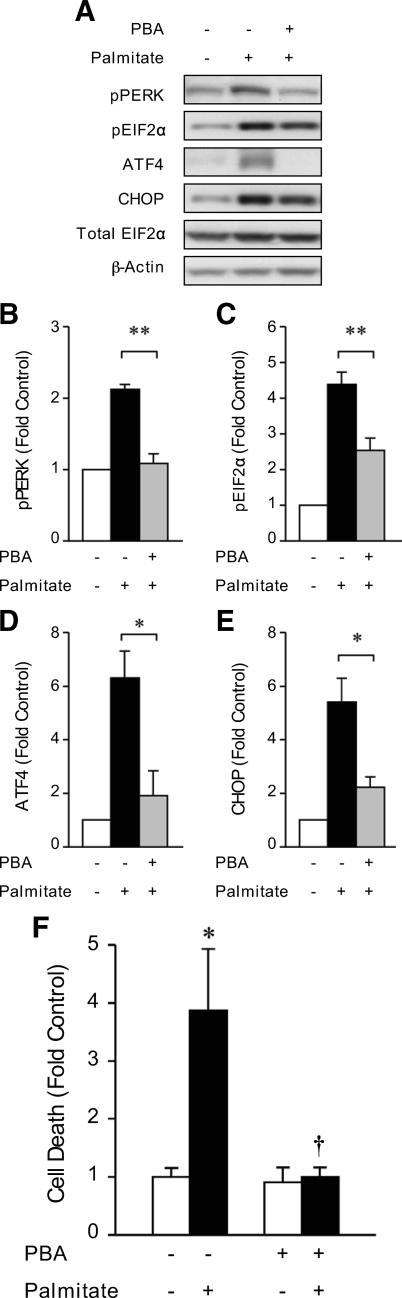
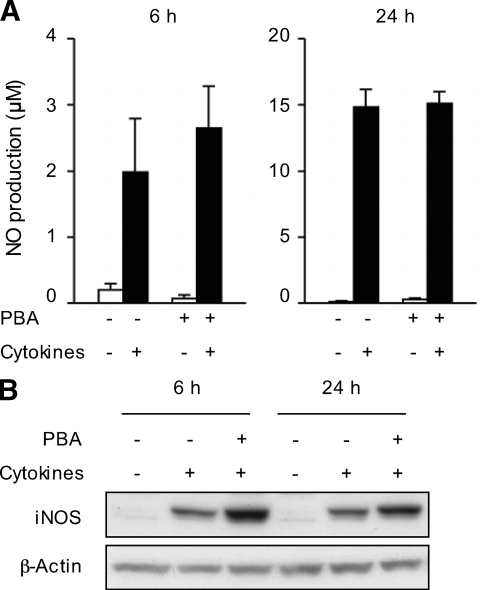
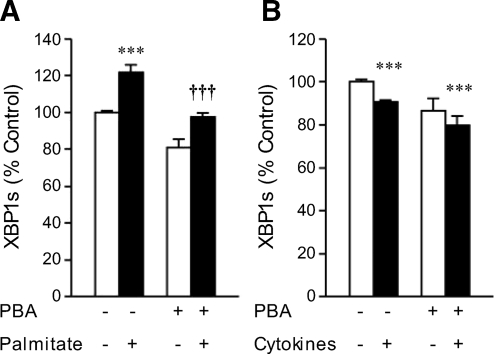
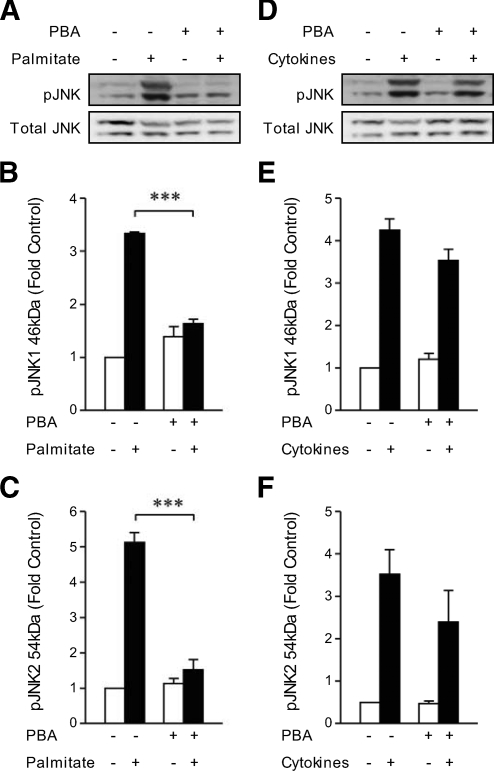
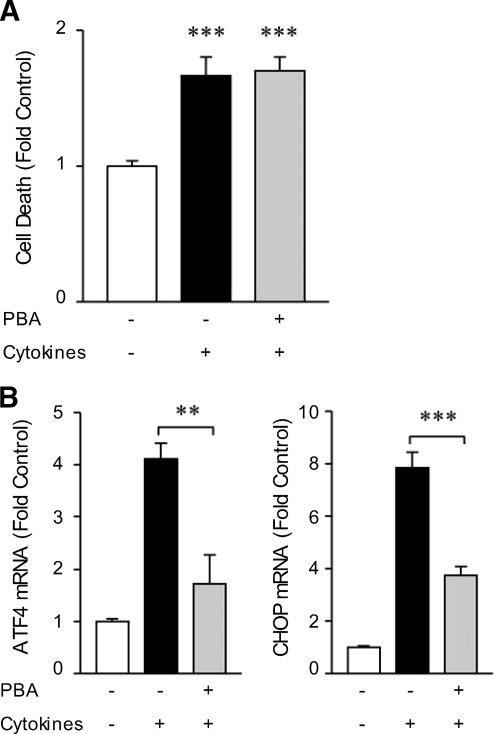
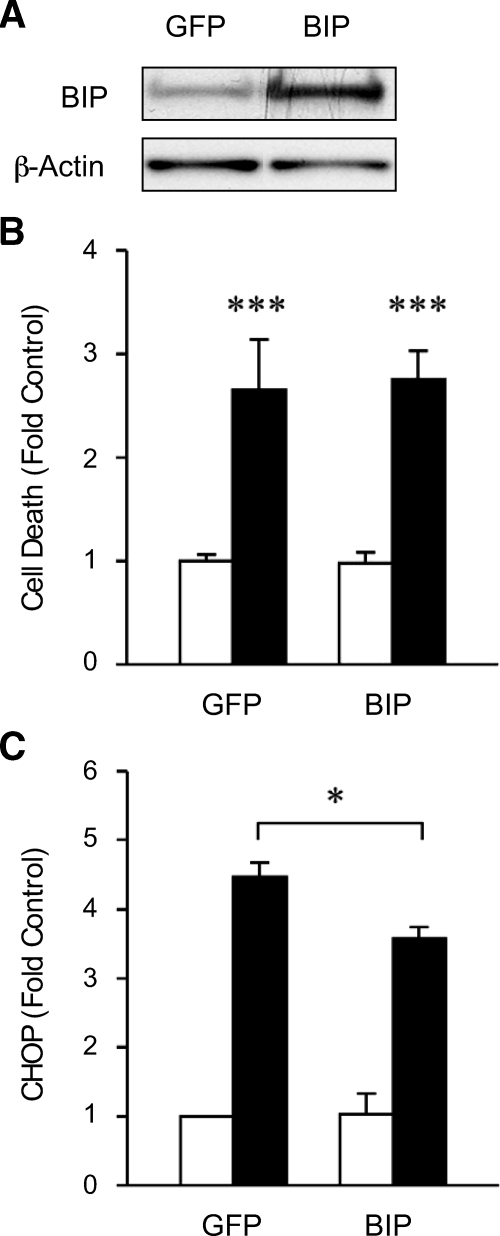
Results: Cytokines and palmitate triggered ER stress in beta-cells as evidenced by increased phosphorylation of PKR-like ER kinase (PERK), eukaryotic initiation factor (EIF)2alpha, and Jun NH(2)-terminal kinase (JNK) and increased expression of activating transcription factor (ATF)4 and CHOP. PBA treatment attenuated ER stress, but JNK phosphorylation was reduced only in response to palmitate, not in response to cytokines. PBA had no effect on cytokine-induced cell death but was associated with protection against palmitate-induced cell death. Similarly, siRNA-mediated reduction in CHOP expression protected against palmitate- but not against cytokine-induced cell death. In NOD islets, mRNA levels of several ER stress genes were reduced (ATF4, BiP [binding protein], GRP94 [glucose regulated protein 94], p58, and XBP-1 [X-box binding protein 1] splicing) or unchanged (CHOP and Edem1 [ER degradation enhancer, mannosidase alpha-like 1]).
Conclusions: While both cytokines and palmitate can induce ER stress, our results suggest that, in contrast to lipoapoptosis, the PERK-ATF4-CHOP ER stress-signaling pathway is not necessary for cytokine-induced beta-cell death.
Figures
References
-
- Eizirik DL, Mandrup-Poulsen T: A choice of death: the signal-transduction of immune-mediated beta-cell apoptosis. Diabetologia 44: 2115–2133, 2001 - PubMed
-
- Rabinovitch A, Suarez-Pinzon WL: Role of cytokines in the pathogenesis of autoimmune diabetes mellitus. Rev Endocr Metab Disord 4: 291–299, 2003 - PubMed
-
- Thomas HE, Darwiche R, Corbett JA, Kay TW: Interleukin-1 plus γ-interferon–induced pancreatic β-cell dysfunction is mediated by β-cell nitric oxide production. Diabetes 51: 311–316, 2002 - PubMed
-
- Cnop M, Welsh N, Jonas JC, Jorns A, Lenzen S, Eizirik DL: Mechanisms of pancreatic β-cell death in type 1 and type 2 diabetes: many differences, few similarities. Diabetes 54: S97–S107, 2005 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
