

Distinct activities of the DExD/H-box splicing factor hUAP56 facilitate stepwise assembly of the spliceosome
- PMID: 18593880
- PMCID: PMC2492666
- DOI: 10.1101/gad.1657308
Distinct activities of the DExD/H-box splicing factor hUAP56 facilitate stepwise assembly of the spliceosome
Abstract
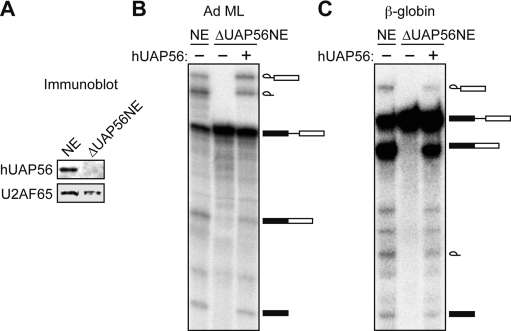
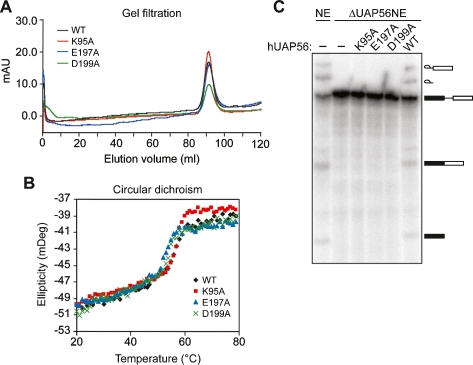
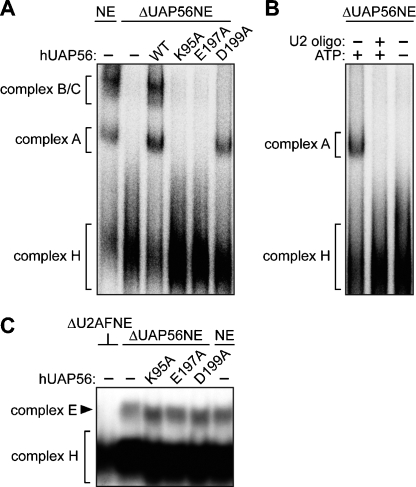
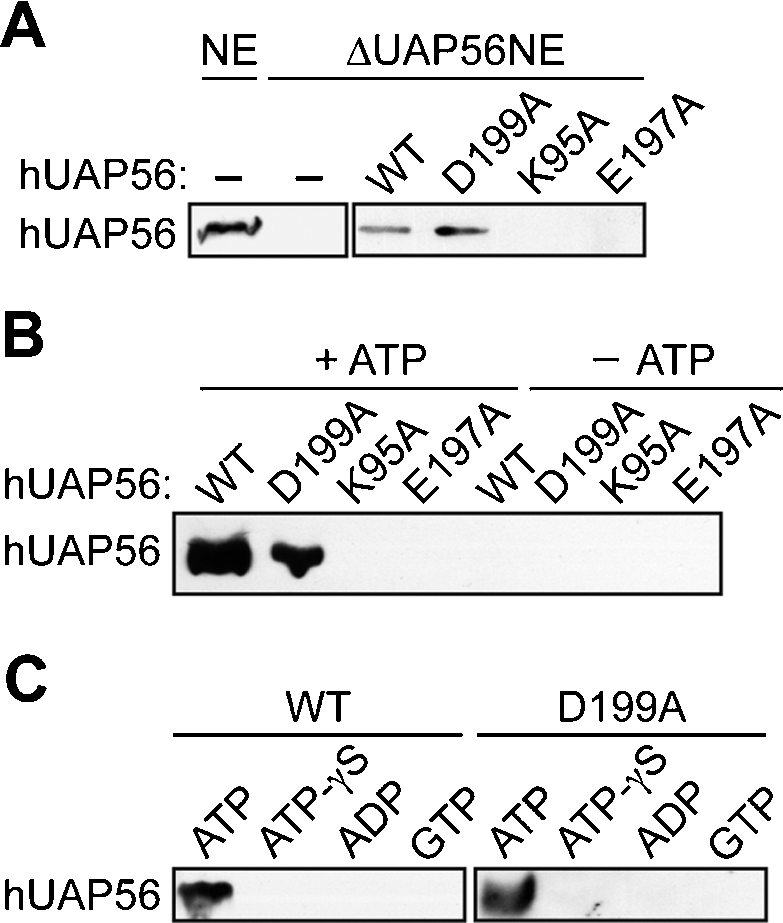
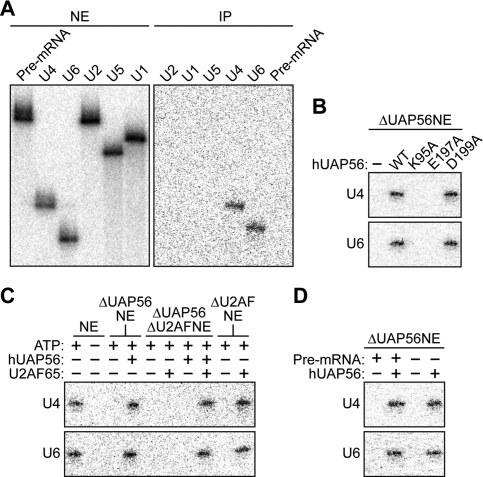
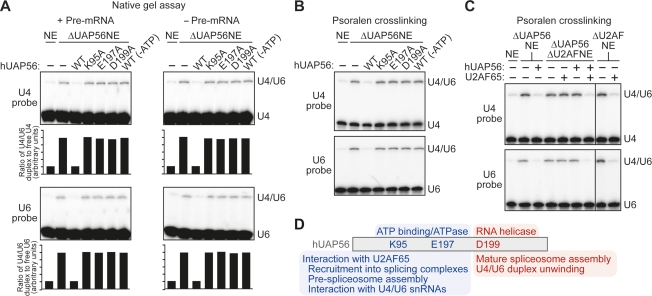
The essential splicing factor human UAP56 (hUAP56) is a DExD/H-box protein known to promote prespliceosome assembly. Here, using a series of hUAP56 mutants that are defective for ATP-binding, ATP hydrolysis, or dsRNA unwindase/helicase activity, we assess the relative contributions of these biochemical functions to pre-mRNA splicing. We show that prespliceosome assembly requires hUAP56's ATP-binding and ATPase activities, which, unexpectedly, are required for hUAP56 to interact with U2AF(65) and be recruited into splicing complexes. Surprisingly, we find that hUAP56 is also required for mature spliceosome assembly, which requires, in addition to the ATP-binding and ATPase activities, hUAP56's dsRNA unwindase/helicase activity. We demonstrate that hUAP56 directly contacts U4 and U6 snRNAs and can promote unwinding of the U4/U6 duplex, and that both these activities are dependent on U2AF(65). Our results indicate that hUAP56 first interacts with U2AF(65) in an ATP-dependent manner, and subsequently with U4/U6 snRNAs to facilitate stepwise assembly of the spliceosome.
Figures
Similar articles
-
RNA unwinding in U4/U6 snRNPs requires ATP hydrolysis and the DEIH-box splicing factor Brr2.Curr Biol. 1998 Jul 16;8(15):847-55. doi: 10.1016/s0960-9822(07)00345-4. Curr Biol. 1998. PMID: 9705931
-
DExD/H-box Prp5 protein is in the spliceosome during most of the splicing cycle.RNA. 2009 Jul;15(7):1345-62. doi: 10.1261/rna.1065209. Epub 2009 May 18. RNA. 2009. PMID: 19451545 Free PMC article.
-
The DEAH-box protein PRP22 is an ATPase that mediates ATP-dependent mRNA release from the spliceosome and unwinds RNA duplexes.EMBO J. 1998 May 15;17(10):2926-37. doi: 10.1093/emboj/17.10.2926. EMBO J. 1998. PMID: 9582286 Free PMC article.
-
Mechanisms of fidelity in pre-mRNA splicing.Curr Opin Cell Biol. 2000 Jun;12(3):340-5. doi: 10.1016/s0955-0674(00)00097-1. Curr Opin Cell Biol. 2000. PMID: 10801464 Review.
-
UAP56- a key player with surprisingly diverse roles in pre-mRNA splicing and nuclear export.BMB Rep. 2009 Apr 30;42(4):185-8. doi: 10.5483/bmbrep.2009.42.4.185. BMB Rep. 2009. PMID: 19403039 Review.
Cited by
-
Host Cell Factors That Interact with Influenza Virus Ribonucleoproteins.Cold Spring Harb Perspect Med. 2021 Nov 1;11(11):a038307. doi: 10.1101/cshperspect.a038307. Cold Spring Harb Perspect Med. 2021. PMID: 32988980 Free PMC article. Review.
-
TREX (transcription/export)-NP complex exerts a dual effect on regulating polymerase activity and replication of influenza A virus.PLoS Pathog. 2022 Sep 9;18(9):e1010835. doi: 10.1371/journal.ppat.1010835. eCollection 2022 Sep. PLoS Pathog. 2022. PMID: 36084138 Free PMC article.
-
The Cellular DExD/H-Box RNA Helicase UAP56 Co-localizes With the Influenza A Virus NS1 Protein.Front Microbiol. 2018 Sep 12;9:2192. doi: 10.3389/fmicb.2018.02192. eCollection 2018. Front Microbiol. 2018. PMID: 30258431 Free PMC article.
-
TRAP150 activates pre-mRNA splicing and promotes nuclear mRNA degradation.Nucleic Acids Res. 2010 Jun;38(10):3340-50. doi: 10.1093/nar/gkq017. Epub 2010 Jan 31. Nucleic Acids Res. 2010. PMID: 20123736 Free PMC article.
-
Stem-loop 4 of U1 snRNA is essential for splicing and interacts with the U2 snRNP-specific SF3A1 protein during spliceosome assembly.Genes Dev. 2014 Nov 15;28(22):2518-31. doi: 10.1101/gad.248625.114. Genes Dev. 2014. PMID: 25403181 Free PMC article.
References
-
- Benz J., Trachsel H., Baumann U. Crystal structure of the ATPase domain of translation initiation factor 4A from Saccharomyces cerevisiae—The prototype of the DEAD box protein family. Structure. 1999;7:671–679. - PubMed
-
- Caruthers J.M., McKay D.B. Helicase structure and mechanism. Curr. Opin. Struct. Biol. 2002;12:123–133. - PubMed
-
- Cordin O., Banroques J., Tanner N.K., Linder P. The DEAD-box protein family of RNA helicases. Gene. 2006;367:17–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases