

The warburg effect in leukemia-stroma cocultures is mediated by mitochondrial uncoupling associated with uncoupling protein 2 activation
- PMID: 18593920
- PMCID: PMC2562568
- DOI: 10.1158/0008-5472.CAN-08-0555
The warburg effect in leukemia-stroma cocultures is mediated by mitochondrial uncoupling associated with uncoupling protein 2 activation
Abstract
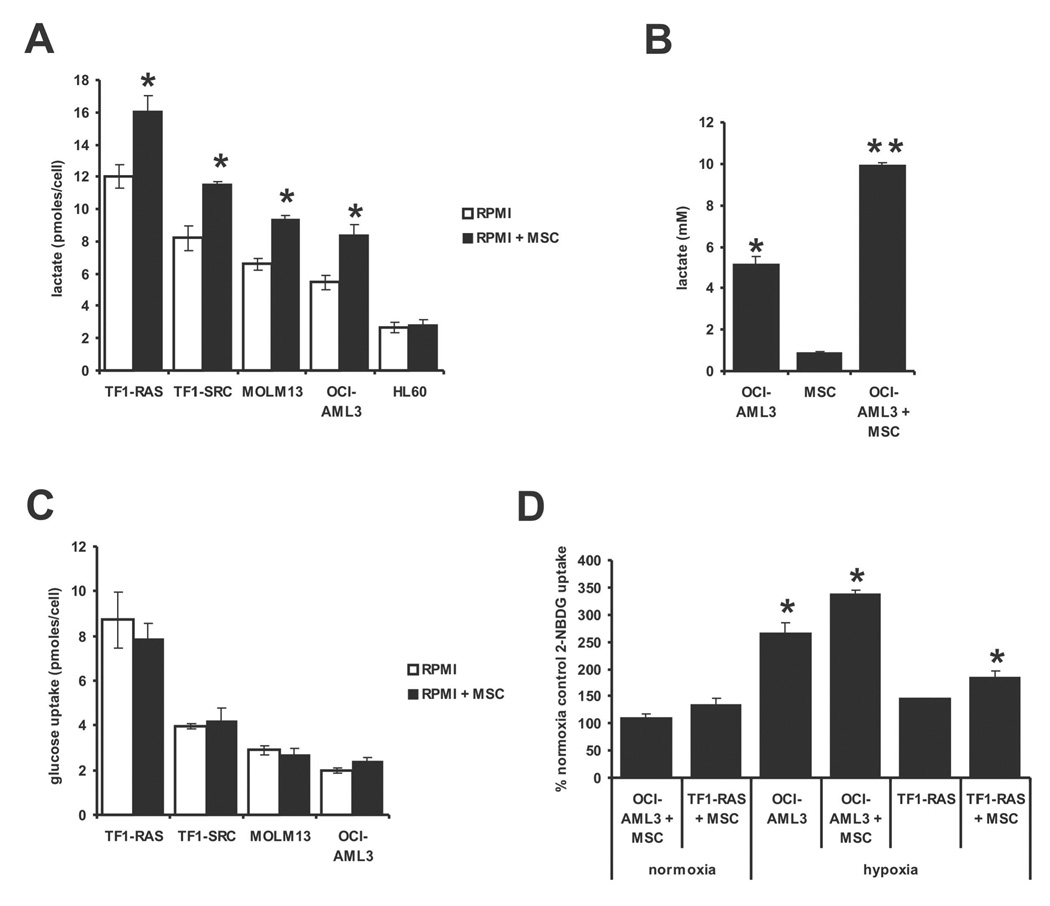
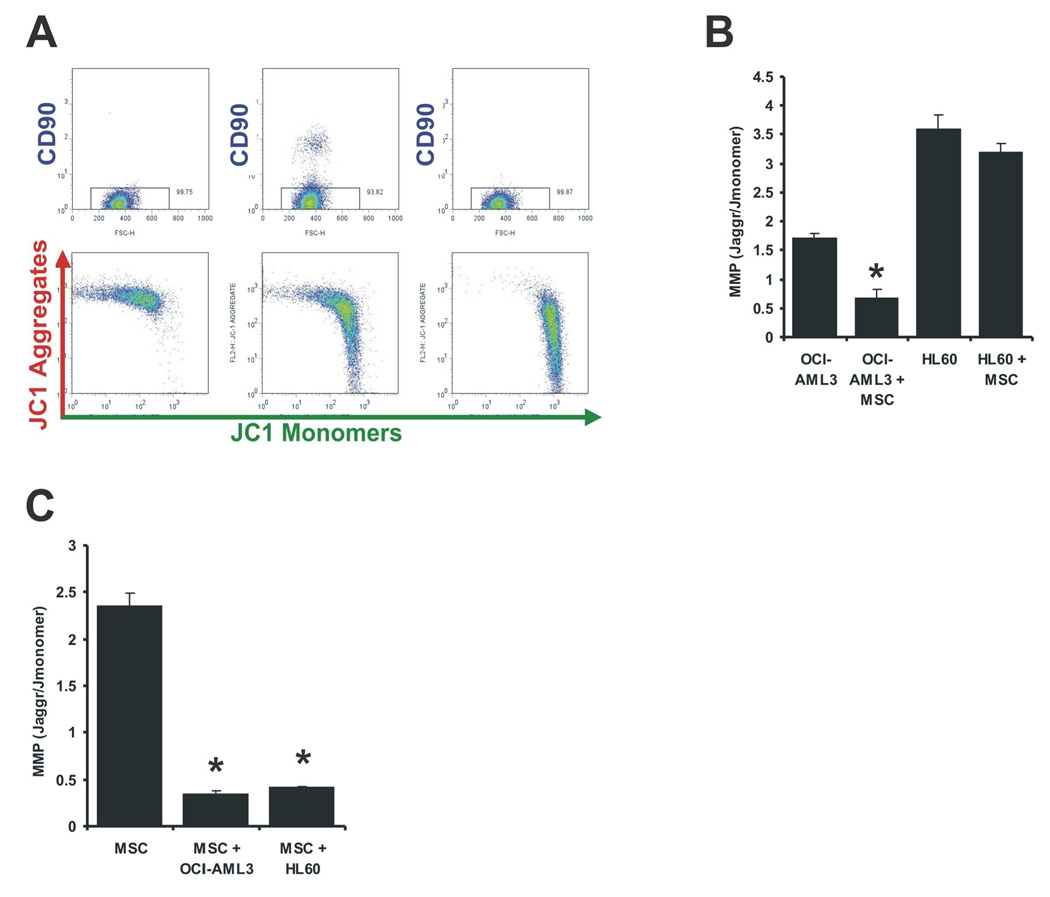
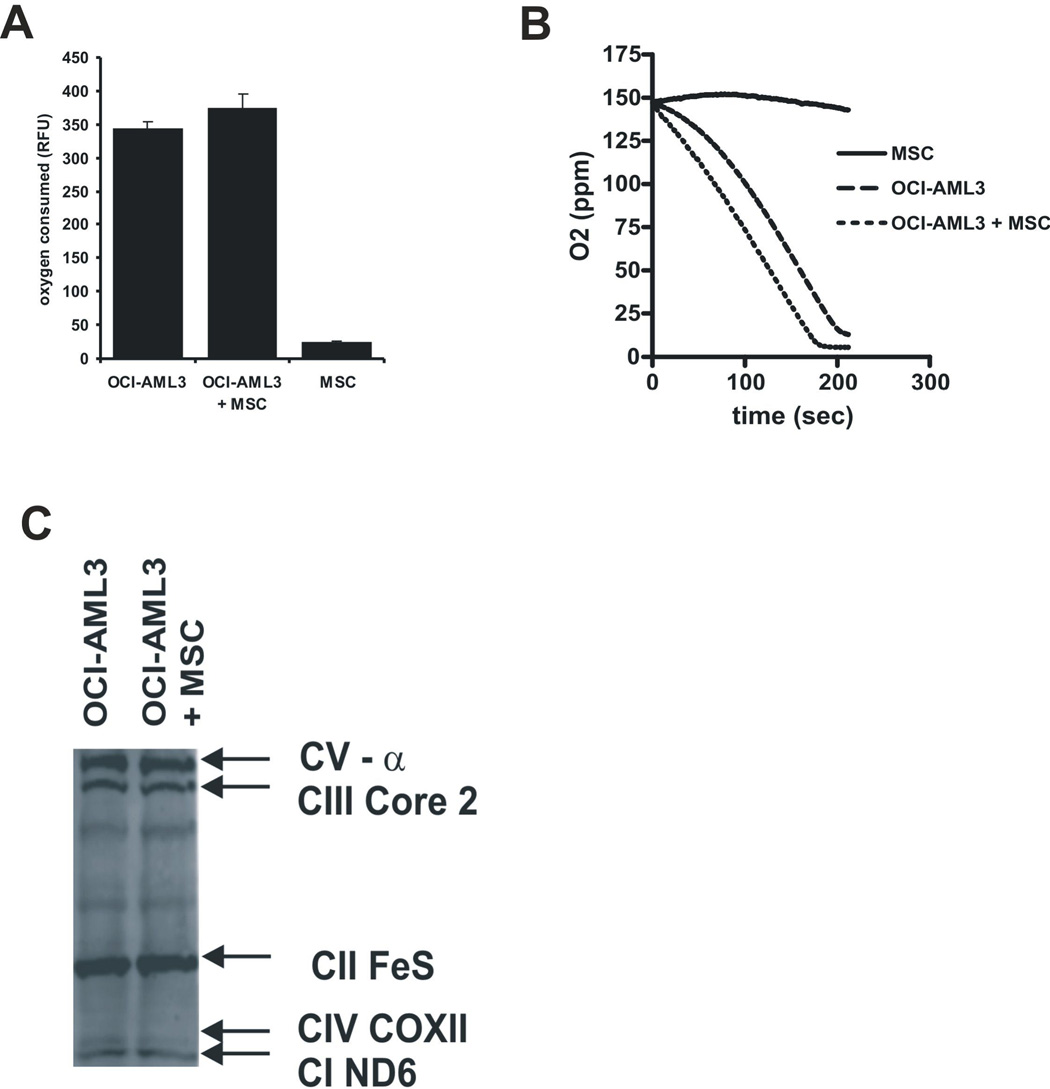
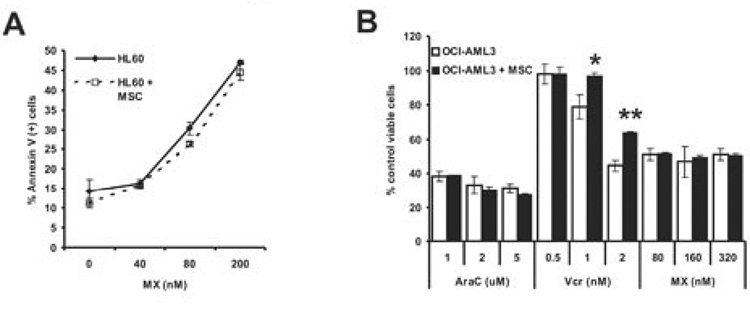
In 1956, Otto Warburg proposed that the origin of cancer cells was closely linked to a permanent respiratory defect that bypassed the Pasteur effect (i.e., the inhibition of anaerobic fermentation by oxygen). Since then, permanent defects in oxygen consumption that could explain the dependence of cancer cells on aerobic glycolysis have not been identified. Here, we show that under normoxic conditions exposure of leukemia cells to bone marrow-derived mesenchymal stromal cells (MSC) promotes accumulation of lactate in the culture medium and reduces mitochondrial membrane potential (DeltaPsiM) in both cell types. Notably, the consumption of glucose was not altered in cocultures, suggesting that the accumulation of lactate was the result of reduced pyruvate metabolism. Interestingly, the decrease in DeltaPsiM was mediated by mitochondrial uncoupling in leukemia cells and was accompanied by increased expression of uncoupling protein 2 (UCP2). HL60 cells fail to increase UCP2 expression, are not uncoupled after coculture, and do not exhibit increased aerobic glycolysis, whereas small interfering RNA-mediated suppression of UCP2 in OCI-AML3 cells reversed mitochondrial uncoupling and aerobic glycolysis elicited by MSC. Taken together, these data suggest that microenvironment activation of highly conserved mammalian UCPs may facilitate the Warburg effect in the absence of permanent respiratory impairment.
Figures



Similar articles
-
Increased activity of mitochondrial uncoupling protein 2 improves stress resistance in cultured endothelial cells exposed in vitro to high glucose levels.Am J Physiol Heart Circ Physiol. 2015 Jul 1;309(1):H147-56. doi: 10.1152/ajpheart.00759.2014. Epub 2015 Apr 24. Am J Physiol Heart Circ Physiol. 2015. PMID: 25910810
-
UCP2, not a physiologically relevant uncoupler but a glucose sparing switch impacting ROS production and glucose sensing.Biochim Biophys Acta. 2009 May;1787(5):377-83. doi: 10.1016/j.bbabio.2009.01.003. Epub 2009 Jan 20. Biochim Biophys Acta. 2009. PMID: 19413946 Review.
-
Cellular model of Warburg effect identifies tumor promoting function of UCP2 in breast cancer and its suppression by genipin.PLoS One. 2011;6(9):e24792. doi: 10.1371/journal.pone.0024792. Epub 2011 Sep 15. PLoS One. 2011. PMID: 21935467 Free PMC article.
-
Uncoupling protein-2 modulates myocardial excitation-contraction coupling.Circ Res. 2010 Mar 5;106(4):730-8. doi: 10.1161/CIRCRESAHA.109.206631. Epub 2010 Jan 7. Circ Res. 2010. PMID: 20056920
-
Mitochondrial recoupling: a novel therapeutic strategy for cancer?Br J Cancer. 2011 Aug 9;105(4):469-74. doi: 10.1038/bjc.2011.245. Epub 2011 Jun 28. Br J Cancer. 2011. PMID: 21712825 Free PMC article. Review.
Cited by
-
Disruption of gap junctions attenuates acute myeloid leukemia chemoresistance induced by bone marrow mesenchymal stromal cells.Oncogene. 2020 Feb;39(6):1198-1212. doi: 10.1038/s41388-019-1069-y. Epub 2019 Oct 24. Oncogene. 2020. PMID: 31649334 Free PMC article.
-
Uncoupling protein 2 and metabolic diseases.Mitochondrion. 2017 May;34:135-140. doi: 10.1016/j.mito.2017.03.005. Epub 2017 Mar 25. Mitochondrion. 2017. PMID: 28351676 Free PMC article. Review.
-
TKTL1 is activated by promoter hypomethylation and contributes to head and neck squamous cell carcinoma carcinogenesis through increased aerobic glycolysis and HIF1alpha stabilization.Clin Cancer Res. 2010 Feb 1;16(3):857-66. doi: 10.1158/1078-0432.CCR-09-2604. Epub 2010 Jan 26. Clin Cancer Res. 2010. PMID: 20103683 Free PMC article.
-
Pharmacologic inhibition of fatty acid oxidation sensitizes human leukemia cells to apoptosis induction.J Clin Invest. 2010 Jan;120(1):142-56. doi: 10.1172/JCI38942. Epub 2009 Dec 21. J Clin Invest. 2010. PMID: 20038799 Free PMC article.
-
Novel Mango Ginger Bioactive (2,4,6-Trihydroxy-3,5-diprenyldihydrochalcone) Inhibits Mitochondrial Metabolism in Combination with Avocatin B.ACS Omega. 2022 Jan 7;7(2):1682-1693. doi: 10.1021/acsomega.1c04053. eCollection 2022 Jan 18. ACS Omega. 2022. PMID: 35071863 Free PMC article.
References
-
- Warburg O. On the origin of cancer cells. Science. 1956;123:309–314. - PubMed
-
- Lynen F. Die Rolle der Phosphorsaeure bei Dehydrierungsovorgaegen und ihre biologische Bedeutung. Die Naturwissenschaften. 1951;30:398.
-
- Ronzoni E, Ehrenfest E. The effect of dinitrophenol on the metabolism of frog muscle. Journal of Biological Chemistry. 1936;15:749.
-
- Desquiret V, Loiseau D, Jacques C, et al. Dinitrophenol-induced mitochondrial uncoupling in vivo triggers respiratory adaptation in HepG2 cells. Biochim Biophys Acta. 2006;1757:21–30. - PubMed
-
- Bouillaud F, Ricquier D, Mory G, Thibault J. Increased level of mRNA for the uncoupling protein in brown adipose tissue of rats during thermogenesis induced by cold exposure or norepinephrine infusion. J Biol Chem. 1984;259:11583–11586. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
