

The dynamic envelope of a fusion class II virus. E3 domain of glycoprotein E2 precursor in Semliki Forest virus provides a unique contact with the fusion protein E1
- PMID: 18596032
- PMCID: PMC3258923
- DOI: 10.1074/jbc.M801470200
The dynamic envelope of a fusion class II virus. E3 domain of glycoprotein E2 precursor in Semliki Forest virus provides a unique contact with the fusion protein E1
Abstract
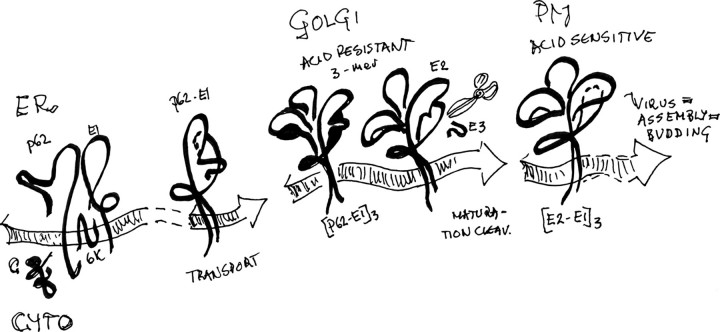
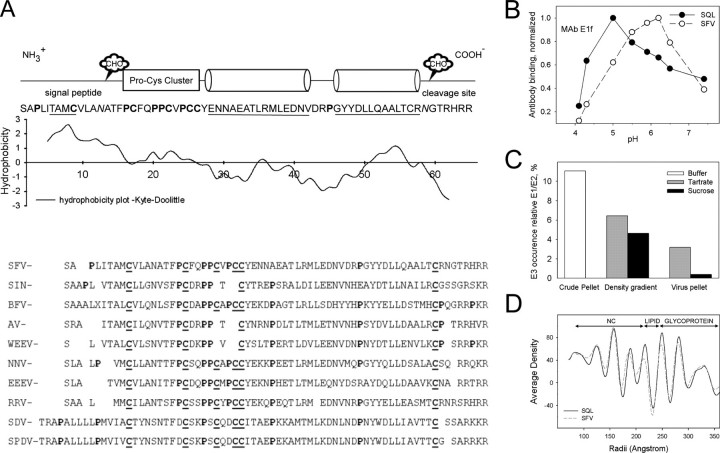
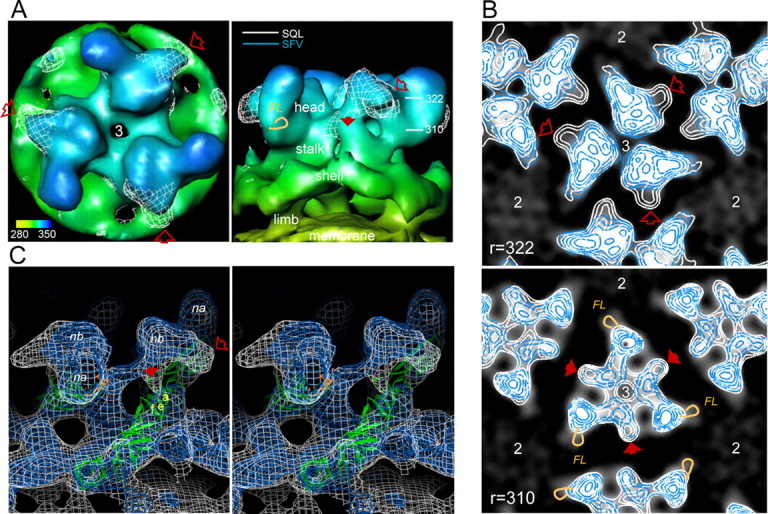
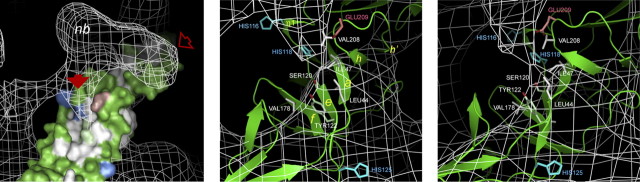
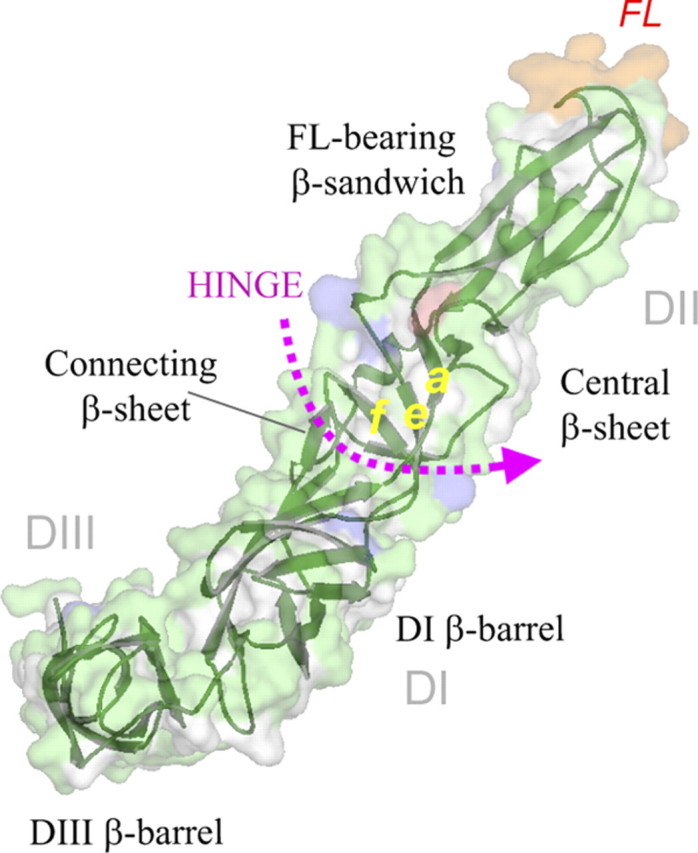
In alphaviruses, here represented by Semliki Forest virus, infection requires an acid-responsive spike configuration to facilitate membrane fusion. The creation of this relies on the chaperone function of glycoprotein E2 precursor (p62) and its maturation cleavage into the small external E3 and the membrane-anchored E2 glycoproteins. To reveal how the E3 domain of p62 exerts its control of spike functions, we determine the structure of a p62 cleavage-impaired mutant virus particle (SQL) by electron cryomicroscopy. A comparison with the earlier solved wild type virus structure reveals that the E3 domain of p62(SQL) forms a bulky side protrusion in the spike head region. This establishes a gripper over part of domain II of the fusion protein, with a cotter-like connection downward to a hydrophobic cluster in its central beta-sheet. This finding reevaluates the role of the precursor from being only a provider of a shield over the fusion loop to a structural playmate in formation of the fusogenic architecture.
Figures
References
-
- Kielian, M. (2006) Virology 344 38–47 - PubMed
-
- White, J. M., Hoffman, L. R., Arevalo, J. H., and Wilson, I. A. (1997) in Structural Biology of Viruses (Chiu, W., Burnett, R. M., and Garcea, R. L., eds) pp. 80–104, Oxford University Press, Oxford
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
