

A proteomics analysis of yeast Mot1p protein-protein associations: insights into mechanism
- PMID: 18596064
- PMCID: PMC2577210
- DOI: 10.1074/mcp.M800221-MCP200
A proteomics analysis of yeast Mot1p protein-protein associations: insights into mechanism
Abstract
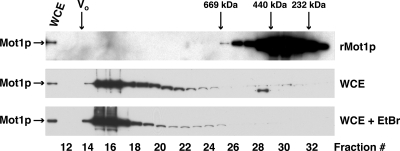
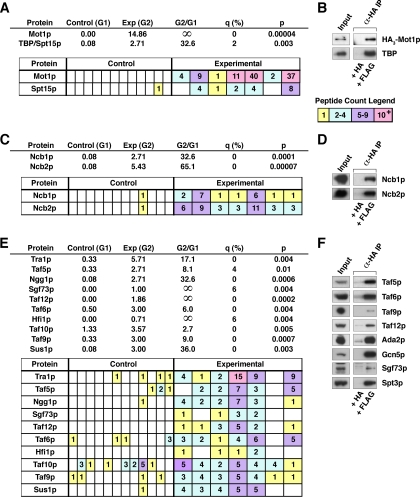
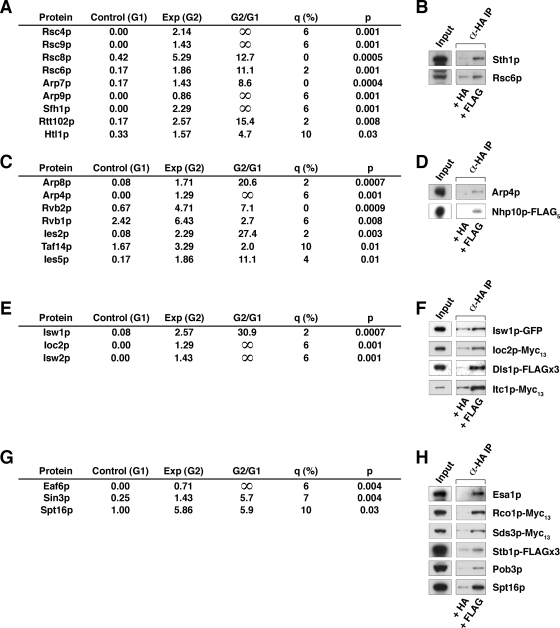
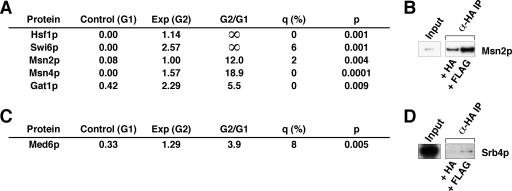
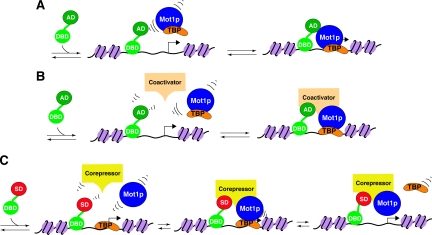
Yeast Mot1p, a member of the Snf2 ATPase family of proteins, is a transcriptional regulator that has the unusual ability to both repress and activate mRNA gene transcription. To identify interactions with other proteins that may assist Mot1p in its regulatory processes, Mot1p was purified from replicate yeast cell extracts, and Mot1p-associated proteins were identified by coupled multidimensional liquid chromatography and tandem mass spectrometry. Using this approach we generated a catalog of Mot1p-interacting proteins. Mot1p interacts with a range of transcriptional co-regulators as well as proteins involved in chromatin remodeling. We propose that interaction with such a wide range of proteins may be one mechanism through which Mot1p subserves its roles as a transcriptional activator and repressor.
Figures
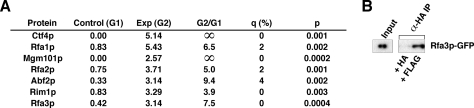
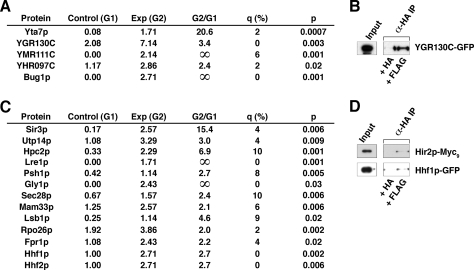
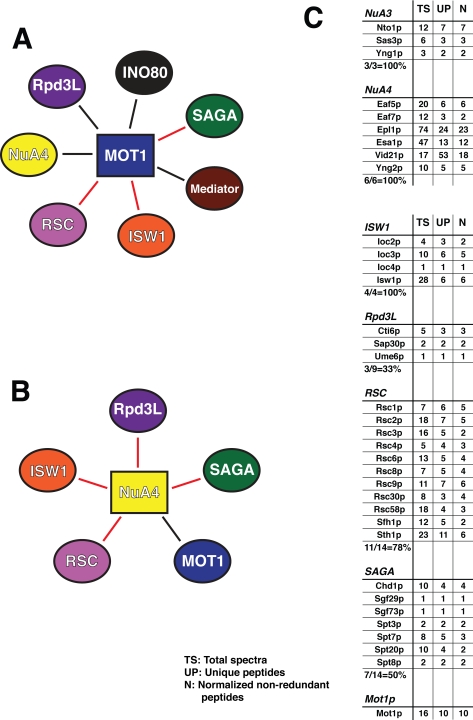
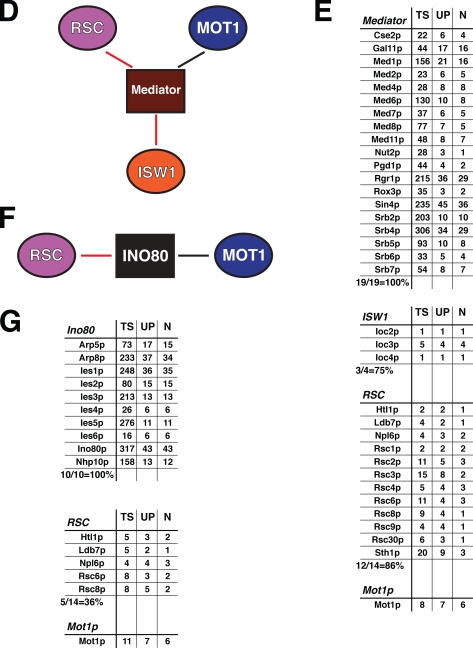
References
-
- Pugh, B. F. ( 2000) Control of gene expression through regulation of the TATA-binding protein. Gene (Amst.) 255, 1–14 - PubMed
-
- Li, X. Y., Virbasius, A., Zhu, X., and Green, M. R. ( 1999) Enhancement of TBP binding by activators and general transcription factors. Nature 399, 605–609 - PubMed
-
- Roeder, R. G. ( 2005) Transcriptional regulation and the role of diverse coactivators in animal cells. FEBS Lett. 579, 909–915 - PubMed
-
- Albright, S. R., and Tjian, R. ( 2000) TAFs revisited: more data reveal new twists and confirm old ideas. Gene (Amst.) 242, 1–13 - PubMed
-
- Lee, T. I., and Young, R. A. ( 1998) Regulation of gene expression by TBP-associated proteins. Genes Dev. 12, 1398–1408 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- N01 AI040079/AI/NIAID NIH HHS/United States
- R01 CA126218/CA/NCI NIH HHS/United States
- P01 HL068744/HL/NHLBI NIH HHS/United States
- P50 CA098131/CA/NCI NIH HHS/United States
- R01 GM052461/GM/NIGMS NIH HHS/United States
- GM64779/GM/NIGMS NIH HHS/United States
- R01 GM064779/GM/NIGMS NIH HHS/United States
- ES11993/ES/NIEHS NIH HHS/United States
- GM52461/GM/NIGMS NIH HHS/United States
- HL68744/HL/NHLBI NIH HHS/United States
- R01 ES011993/ES/NIEHS NIH HHS/United States
- HHSN266200400079C/PHS HHS/United States
- R56 GM052461/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
