

P2X7 receptor-Pannexin1 complex: pharmacology and signaling
- PMID: 18596211
- PMCID: PMC2544446
- DOI: 10.1152/ajpcell.00228.2008
P2X7 receptor-Pannexin1 complex: pharmacology and signaling
Erratum in
- Am J Physiol Cell Physiol. 2009 Mar;296(3):C639
Abstract
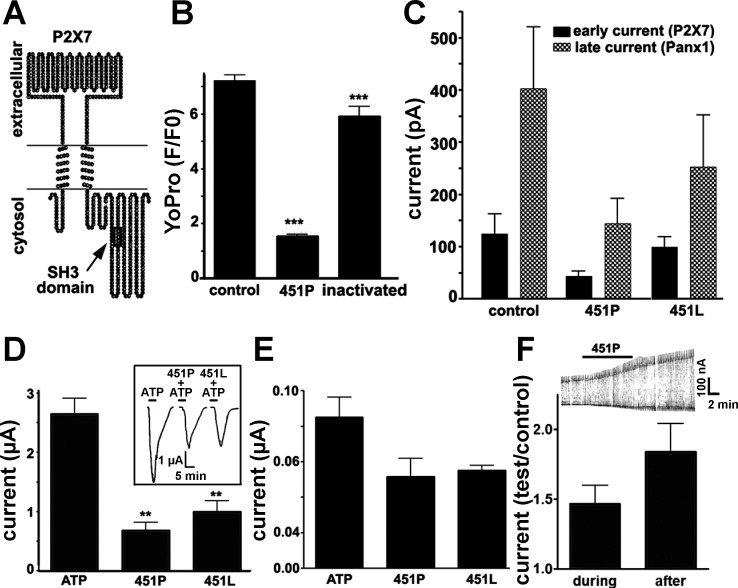
Pannexin 1 (Panx1), an ortholog to invertebrate innexin gap junctions, has recently been proposed to be the pore induced by P2X(7) receptor (P2X(7)R) activation. We explored the pharmacological action of compounds known to block gap junctions on Panx1 channels activated by the P2X(7)R and the mechanisms involved in the interaction between these two proteins. Whole cell recordings revealed distinct P2X(7)R and Panx1 currents in response to agonists. Activation of Panx1 currents following P2X(7)R stimulation or by membrane depolarization was blocked by Panx1 small-interfering RNA (siRNA) and with mefloquine > carbenoxolone > flufenamic acid. Incubation of cells with KN-62, a P2X(7)R antagonist, prevented current activation by 2'(3')-O-(4-benzoylbenzoyl)adenosine 5'-triphosphate (BzATP). Membrane permeabilization to dye induced by BzATP was also prevented by Panx1 siRNA and by carbenoxolone and mefloquine. Membrane permeant (TAT-P2X(7)) peptides, provided evidence that the Src homology 3 death domain of the COOH-terminus of the P2X(7)R is involved in the initial steps of the signal transduction events leading to Panx1 activation and that a Src tyrosine kinase is likely involved in this process. Competition assays indicated that 20 microM TAT-P2X(7) peptide caused 50% reduction in Src binding to the P2X(7)R complex. Src tyrosine phosphorylation following BzATP stimulation was reduced by KN-62, TAT-P2X(7) peptide, and by the Src tyrosine inhibitor PP2 and these compounds prevented both large-conductance Panx1 currents and membrane permeabilization. These results together with the lack Panx1 tyrosine phosphorylation in response to P2X(7)R stimulation indicate the involvement of an additional molecule in the tyrosine kinase signal transduction pathway mediating Panx1 activation through the P2X(7)R.
Figures



References
-
- Adriouch S, Dox C, Welge V, Seman M, Koch-Nolte F, Haag F. Cutting edge: a natural P451L mutation in the cytoplasmic domain impairs the function of the mouse P2X7 receptor. J Immunol 169: 4108–4112, 2002. - PubMed
-
- Alves LA, Coutinho-Silva R, Persechini PM, Spray DC, Savino W, Campos de Carvalho AC. Are there functional gap junctions or junctional hemichannels in macrophages? Blood 88: 328–334, 1996. - PubMed
-
- Bao L, Locovei S, Dahl G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett 572: 65–68, 2004. - PubMed
-
- Baraldi PG, Makaeva R, Pavani MG, Nunez MC, Spalluto G, Moro S, Falzoni D, Di Virgilio F, Romagnoli R. Synthesis, biological activity and molecular modeling studies of 1,2,3,4-tetrahydroisoquinoline derivatives as conformationally constrained analogues of KN62, a potent antagonist of the P2X7-receptor containing a tyrosine moiety. Arzeimittelforschung 52: 273–285, 2002. - PubMed
-
- Baranova A, Ivanov D, Petrash N, Pestova A, Skoblov M, Kelmanson I, Shagin D, Nazarenko S, Geraymovych E, Litvin O, Tiunova A, Born TL, Usman N, Staroverov D, Lukyanov S, Panchin Y. The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 83: 706–716, 2004. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous