

On the extent and origins of genic novelty in the phylum Nematoda
- PMID: 18596977
- PMCID: PMC2432500
- DOI: 10.1371/journal.pntd.0000258
On the extent and origins of genic novelty in the phylum Nematoda
Abstract
Background: The phylum Nematoda is biologically diverse, including parasites of plants and animals as well as free-living taxa. Underpinning this diversity will be commensurate diversity in expressed genes, including gene sets associated specifically with evolution of parasitism.
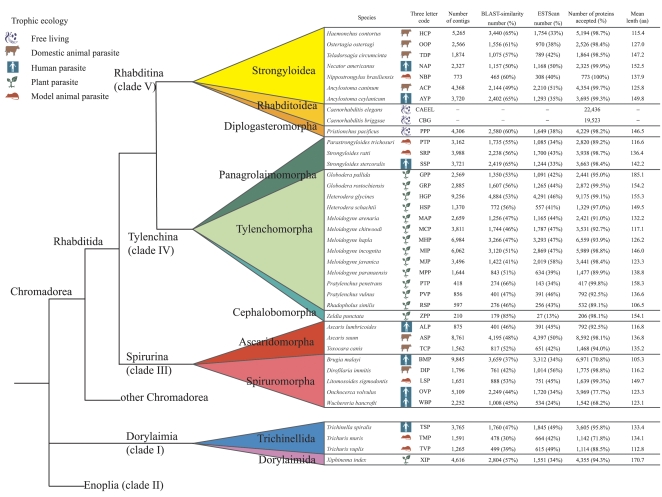
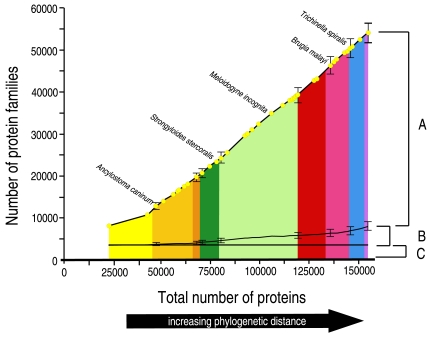
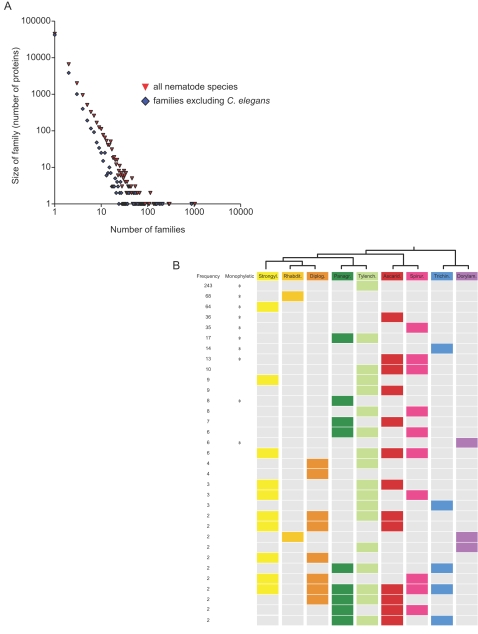
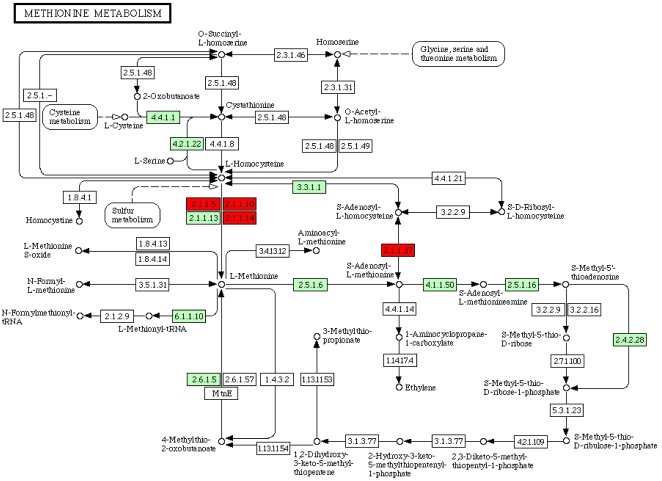
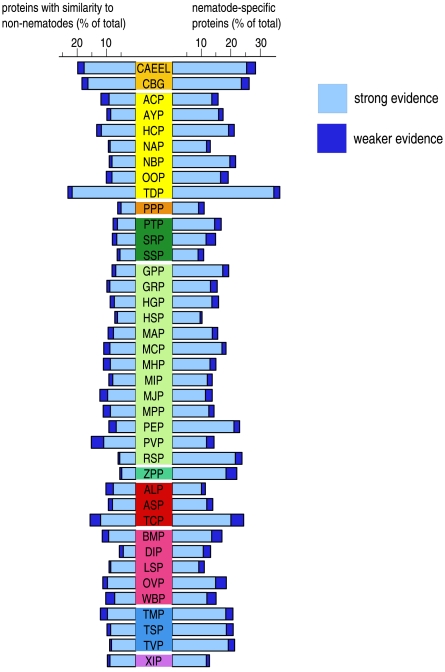
Methods and findings: Here we have analyzed the extensive expressed sequence tag data (available for 37 nematode species, most of which are parasites) and define over 120,000 distinct putative genes from which we have derived robust protein translations. Combined with the complete proteomes of Caenorhabditis elegans and Caenorhabditis briggsae, these proteins have been grouped into 65,000 protein families that in turn contain 40,000 distinct protein domains. We have mapped the occurrence of domains and families across the Nematoda and compared the nematode data to that available for other phyla. Gene loss is common, and in particular we identify nearly 5,000 genes that may have been lost from the lineage leading to the model nematode C. elegans. We find a preponderance of novelty, including 56,000 nematode-restricted protein families and 26,000 nematode-restricted domains. Mapping of the latest time-of-origin of these new families and domains across the nematode phylogeny revealed ongoing evolution of novelty. A number of genes from parasitic species had signatures of horizontal transfer from their host organisms, and parasitic species had a greater proportion of novel, secreted proteins than did free-living ones.
Conclusions: These classes of genes may underpin parasitic phenotypes, and thus may be targets for development of effective control measures.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A transcriptomic analysis of the phylum Nematoda.Nat Genet. 2004 Dec;36(12):1259-67. doi: 10.1038/ng1472. Epub 2004 Nov 14. Nat Genet. 2004. PMID: 15543149
-
Caenorhabditis elegans is a nematode.Science. 1998 Dec 11;282(5396):2041-6. doi: 10.1126/science.282.5396.2041. Science. 1998. PMID: 9851921 Review.
-
Evolution of dnmt-2 and mbd-2-like genes in the free-living nematodes Pristionchus pacificus, Caenorhabditis elegans and Caenorhabditis briggsae.Nucleic Acids Res. 2004 Dec 2;32(21):6388-96. doi: 10.1093/nar/gkh982. Print 2004. Nucleic Acids Res. 2004. PMID: 15576683 Free PMC article.
-
Codon usage patterns in Nematoda: analysis based on over 25 million codons in thirty-two species.Genome Biol. 2006;7(8):R75. doi: 10.1186/gb-2006-7-8-r75. Genome Biol. 2006. PMID: 26271136 Free PMC article.
-
Nematoda: genes, genomes and the evolution of parasitism.Adv Parasitol. 2003;54:101-95. doi: 10.1016/s0065-308x(03)54003-9. Adv Parasitol. 2003. PMID: 14711085 Review.
Cited by
-
s-Adenosylmethionine Levels Govern Innate Immunity through Distinct Methylation-Dependent Pathways.Cell Metab. 2015 Oct 6;22(4):633-45. doi: 10.1016/j.cmet.2015.07.013. Epub 2015 Aug 27. Cell Metab. 2015. PMID: 26321661 Free PMC article.
-
The draft genome of the parasitic nematode Trichinella spiralis.Nat Genet. 2011 Mar;43(3):228-35. doi: 10.1038/ng.769. Epub 2011 Feb 20. Nat Genet. 2011. PMID: 21336279 Free PMC article.
-
Duplication and divergence: the evolution of nematode globins.J Nematol. 2009 Mar;41(1):35-51. J Nematol. 2009. PMID: 22661776 Free PMC article.
-
The genome of a subterrestrial nematode reveals adaptations to heat.Nat Commun. 2019 Nov 21;10(1):5268. doi: 10.1038/s41467-019-13245-8. Nat Commun. 2019. PMID: 31754114 Free PMC article.
-
Co-evolution between an endosymbiont and its nematode host: Wolbachia asymmetric posterior localization and AP polarity establishment.PLoS Negl Trop Dis. 2014 Aug 28;8(8):e3096. doi: 10.1371/journal.pntd.0003096. eCollection 2014 Aug. PLoS Negl Trop Dis. 2014. PMID: 25165813 Free PMC article.
References
-
- Lambshead PJ. Recent developments in marine benthic biodiversity research. Oceanis. 1993;19:5–24.
-
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, et al. The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. - PubMed
-
- Holt RA, Subramanian GM, Halpern A, Sutton GG, Charlab R, et al. The genome sequence of the malaria mosquito Anopheles gambiae. Science. 2002;298:129–149. - PubMed
-
- Mita K, Kasahara M, Sasaki S, Nagayasu Y, Yamada T, et al. The genome sequence of silkworm, Bombyx mori. DNA Res. 2004;11:27–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
