

FUS-DDIT3 prevents the development of adipocytic precursors in liposarcoma by repressing PPARgamma and C/EBPalpha and activating eIF4E
- PMID: 18596980
- PMCID: PMC2434200
- DOI: 10.1371/journal.pone.0002569
FUS-DDIT3 prevents the development of adipocytic precursors in liposarcoma by repressing PPARgamma and C/EBPalpha and activating eIF4E
Abstract
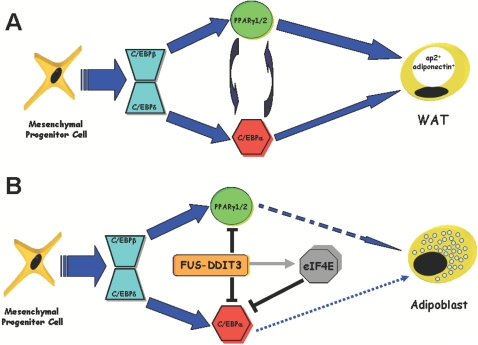
Background: FUS-DDIT3 is a chimeric protein generated by the most common chromosomal translocation t(12;16)(q13;p11) linked to liposarcomas, which are characterized by the accumulation of early adipocytic precursors. Current studies indicate that FUS-DDIT3- liposarcoma develops from uncommitted progenitors. However, the precise mechanism whereby FUS-DDIT3 contributes to the differentiation arrest remains to be elucidated.
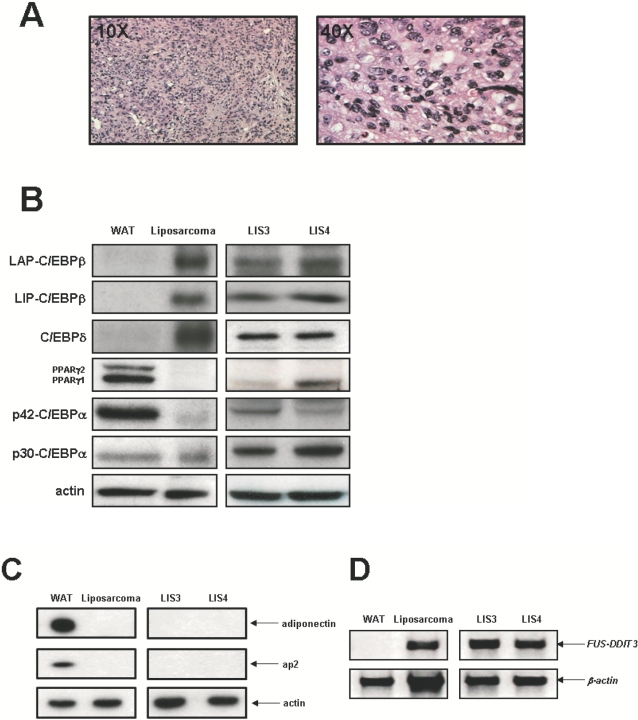
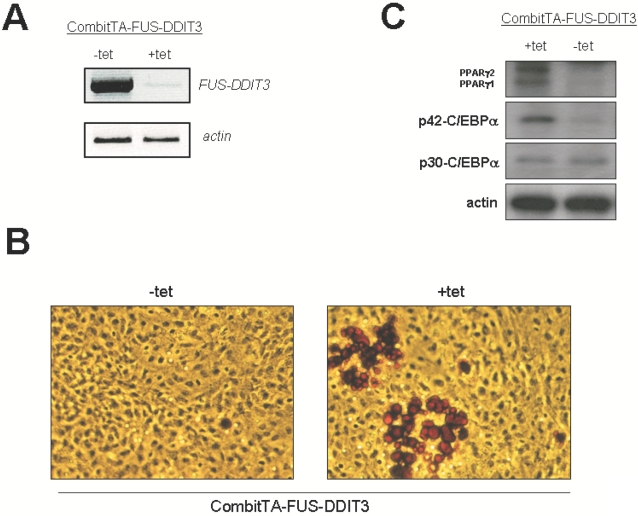
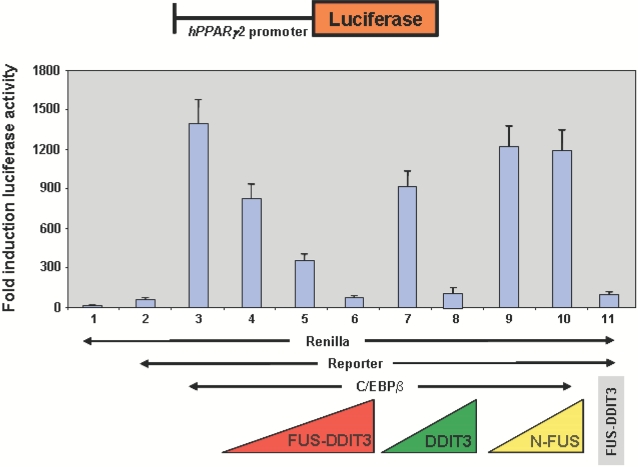
Methodology/principal findings: Here we have characterized the adipocyte regulatory protein network in liposarcomas of FUS-DITT3 transgenic mice and showed that PPARgamma2 and C/EBPalpha expression was altered. Consistent with in vivo data, FUS-DDIT3 MEFs and human liposarcoma cell lines showed a similar downregulation of both PPARgamma2 and C/EBPalpha expression. Complementation studies with PPARgamma but not C/EBPalpha rescued the differentiation block in committed adipocytic precursors expressing FUS-DDIT3. Our results further show that FUS-DDIT3 interferes with the control of initiation of translation by upregulation of the eukaryotic translation initiation factors eIF2 and eIF4E both in FUS-DDIT3 mice and human liposarcomas cell lines, explaining the shift towards the truncated p30 isoform of C/EBPalpha in liposarcomas. Suppression of the FUS-DDIT3 transgene did rescue this adipocyte differentiation block. Moreover, eIF4E was also strongly upregulated in normal adipose tissue of FUS-DDIT3 transgenic mice, suggesting that overexpression of eIF4E may be a primary event in the initiation of liposarcomas. Reporter assays showed FUS-DDIT3 is involved in the upregulation of eIF4E in liposarcomas and that both domains of the fusion protein are required for affecting eIF4E expression.
Conclusions/significance: Taken together, this study provides evidence of the molecular mechanisms involve in the disruption of normal adipocyte differentiation program in liposarcoma harbouring the chimeric gene FUS-DDIT3.
Conflict of interest statement
Figures



Similar articles
-
FUS::DDIT3 Fusion Protein in the Development of Myxoid Liposarcoma and Possible Implications for Therapy.Biomolecules. 2024 Oct 14;14(10):1297. doi: 10.3390/biom14101297. Biomolecules. 2024. PMID: 39456230 Free PMC article. Review.
-
Fat-specific FUS-DDIT3-transgenic mice establish PPARgamma inactivation is required to liposarcoma development.Carcinogenesis. 2007 Oct;28(10):2069-73. doi: 10.1093/carcin/bgm107. Epub 2007 Apr 29. Carcinogenesis. 2007. PMID: 17468515
-
Myxoid liposarcoma-associated EWSR1-DDIT3 selectively represses osteoblastic and chondrocytic transcription in multipotent mesenchymal cells.PLoS One. 2012;7(5):e36682. doi: 10.1371/journal.pone.0036682. Epub 2012 May 3. PLoS One. 2012. PMID: 22570737 Free PMC article.
-
Liposarcoma initiated by FUS/TLS-CHOP: the FUS/TLS domain plays a critical role in the pathogenesis of liposarcoma.Oncogene. 2000 Dec 7;19(52):6015-22. doi: 10.1038/sj.onc.1204018. Oncogene. 2000. PMID: 11146553
-
Understanding mesenchymal cancer: the liposarcoma-associated FUS-DDIT3 fusion gene as a model.Semin Cancer Biol. 2005 Jun;15(3):206-14. doi: 10.1016/j.semcancer.2005.01.006. Semin Cancer Biol. 2005. PMID: 15826835 Review.
Cited by
-
FUS::DDIT3 Fusion Protein in the Development of Myxoid Liposarcoma and Possible Implications for Therapy.Biomolecules. 2024 Oct 14;14(10):1297. doi: 10.3390/biom14101297. Biomolecules. 2024. PMID: 39456230 Free PMC article. Review.
-
Suppression of antitumor cytokine IL‑24 by PRG4 and PAI‑1 may promote myxoid liposarcoma cell survival.Biomed Rep. 2023 Jul 26;19(3):60. doi: 10.3892/br.2023.1642. eCollection 2023 Sep. Biomed Rep. 2023. PMID: 37614985 Free PMC article.
-
Establishment and characterization of a new human myxoid liposarcoma cell line (DL-221) with the FUS-DDIT3 translocation.Lab Invest. 2016 Aug;96(8):885-94. doi: 10.1038/labinvest.2016.64. Epub 2016 Jun 6. Lab Invest. 2016. PMID: 27270875 Free PMC article.
-
Extensive lipoma-like changes of myxoid liposarcoma: morphologic, immunohistochemical, and molecular cytogenetic analyses.Virchows Arch. 2015 Apr;466(4):453-64. doi: 10.1007/s00428-015-1721-z. Epub 2015 Feb 4. Virchows Arch. 2015. PMID: 25650275 Free PMC article.
-
The TET family of proteins: functions and roles in disease.J Mol Cell Biol. 2009 Dec;1(2):82-92. doi: 10.1093/jmcb/mjp025. Epub 2009 Sep 24. J Mol Cell Biol. 2009. PMID: 19783543 Free PMC article. Review.
References
-
- US Cancer Statistics Working Group. Department of Health and Human Services United States Cancer Statistics: 1999 Incidence (Centers for Disease Control and Prevention and National Cancer Institute, Atlanta, 2002)
-
- Mack TM. Sarcomas and other malignancies of soft tissue, retroperitoneum, peritoneum, pleura, heart, mediastinum, and spleen. Cancer. 1995;75:211–244. - PubMed
-
- Aman P, Ron D, Mandahl N, Fioretos T, Heim S, et al. Rearrangement of the transcription factor gene CHOP in myxoid liposarcomas with t(12;16)(q13;p11). Genes Chromosomes Cancer. 1992;5(4):278–285. - PubMed
-
- Crozat A, Aman P, Mandahl N, Ron D. Fusion of CHOP to a novel RNA-binding protein in human myxoid liposarcoma. Nature. 1993;363(6430):640–644. - PubMed
-
- Rabbitts TH, Forster A, Larson R, Nathan P. Fusion of the dominant negative transcription regulator CHOP with a novel gene FUS by translocation t(12;16) in malignant liposarcoma. Nat Genet. 1993;4(2):175–180. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
