

Review
doi: 10.1007/s00418-008-0461-4.
Epub 2008 Jul 3.
Reactive oxygen species in phagocytic leukocytes
Affiliations
- PMID: 18597105
- PMCID: PMC2491708
- DOI: 10.1007/s00418-008-0461-4
Item in Clipboard
Review
Reactive oxygen species in phagocytic leukocytes
Histochem Cell Biol.
2008 Aug.
Abstract
Phagocytic leukocytes consume oxygen and generate reactive oxygen species in response to appropriate stimuli. The phagocyte NADPH oxidase, a multiprotein complex, existing in the dissociated state in resting cells becomes assembled into the functional oxidase complex upon stimulation and then generates superoxide anions. Biochemical aspects of the NADPH oxidase are briefly discussed in this review; however, the major focus relates to the contributions of various modes of microscopy to our understanding of the NADPH oxidase and the cell biology of phagocytic leukocytes.
Figures
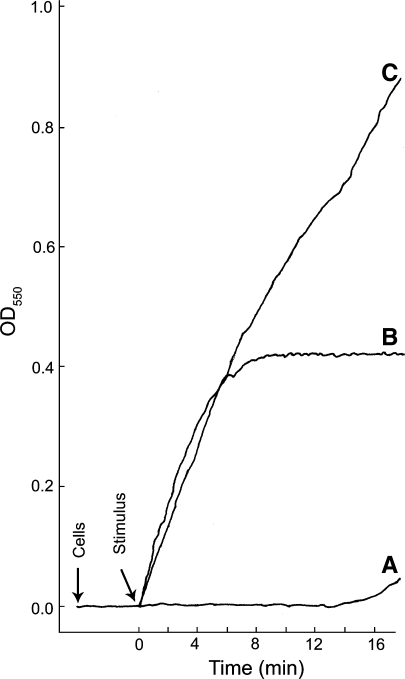
The levels of O2- produced and the rate at which it is produced can vary depending upon the stimulus. Real-time kinetics of O2− production by thioglycolate-elicited guinea pig neutrophils stimulated in three different ways is shown; the cytochrome c reduction spectrophotometric assay was employed. Representative curves of cells stimulated with either (A) NaF (12.5 mM) plus AlCl3 (100 μM), (B) the chemotatic peptide f-Met-Leu-Phe (10−7 M), or (C) a combination of f-Met-Leu-Phe and NaF plus AlCl3 at the same concentrations used in curves A and B. Cells (1 × 106) were added and allowed to equilibrate at 37°C for 3 min before addition of the stimulus; during that time the resting cells did not generate O2−. Following the equilibration period, a stimulus was added (arrow). With NaF plus AlCl3 there was a long lag period prior to the release of modest amounts of O2− (curve A). This lag period is likely due to the slow entry of the stimulus into the cells. The lag period was eliminated when cells were permeabilized by electroporation in the presence of NaF plus AlCl3 (Hartfield and Robinson 1990). When cells were stimulated with the chemotatic peptide, production of O2− was essentially instantaneous and was largely shut down by 5 min post stimulation (curve B). When cells were stimulated by a combination of f-Met-Leu-Phe and NaF plus AlCl3, at the same concentrations used in curves A and B, the initial rate of O2− production was essentially the same as that for the chemotatic peptide alone; however, the shut off of the f-Met-Leu-Phe stimulated O2− was abrogated and the cells continued to produce O2− at a high rate throughout the course of the incubation period. This experiment shows that: (1) resting neutrophils in suspension do not generate O2−; (2) activation of the NADPH oxidase and the production of O2− can be very rapid in intact neutrophils; and (3) different stimulatory conditions can lead to different levels of O2− production and the kinetics of O2− production vary with different stimulatory conditions. The continuous spectrophotometric assay for O2− production was as we have described previously (Robinson et al. 1987)
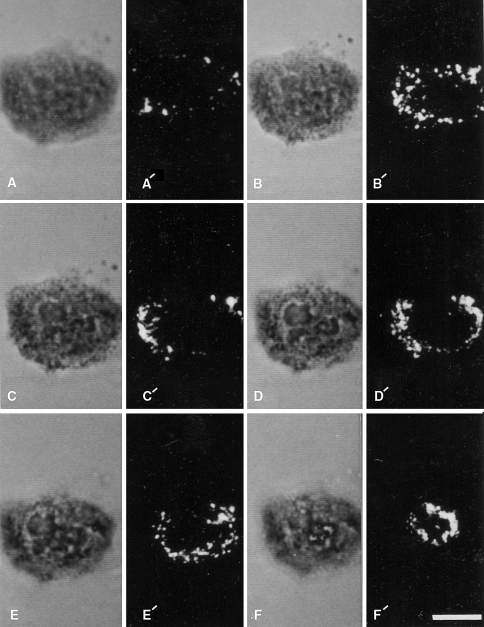
Cytochemical detection of sites of stimulated O2− generation with the NBT reaction and laser scanning confocal microscopy. A single neutrophil was allowed to adhere to a glass cover slip and was subsequently stimulated with 4 β-phorbol 12-myristate acetate in the presence of NBT is shown. The cell was then optically sectioned and viewed by transmitted light (a–f) and by confocal laser scanning microscopy in the reflectance mode (a′–f′). The non-confocal transmitted light images go from the substratum region (a) toward the apical portion of the cell (f). The reflectance images represent confocal sections corresponding to these different focal planes in the transmitted light images. Reaction product is present over the entire cell: however, there is heterogeneity in its distribution. Bar: 10 μm. Reprinted from Robinson and Badwey 1995
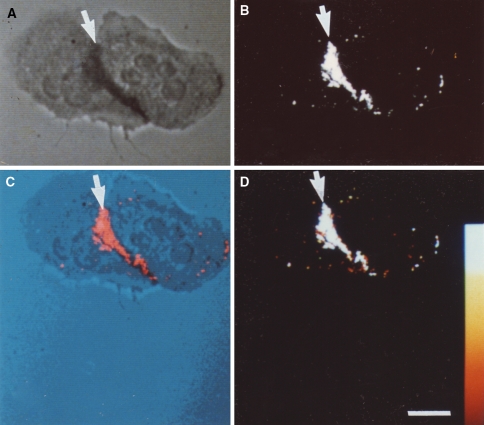
Cytochemical detection of NBT-derived diformazan reaction product by reflected-light laser scanning confocal microscopy. Two neutrophils that were incubated with NBT in the absence of exogenous stimulating agents are shown. a Cells were viewed by non-confocal transmitted light. b The same cells were imaged by laser scanning confocal microscopy in the reflectance mode. The adherent neutrophils have a region of cell–cell contact. The reaction product, indicative of O2− production, is detected in both the transmitted light image and the confocal reflectance image (arrows). c The co-localization of the transmitted light image and the confocal reflectance image is the pseudocolor merge of the two images; the reaction product is indicated by the arrow. d A color wedge has been inserted to estimate the intensity of the reflectance signal. Note that the diformazan reaction product is almost entirely restricted to the area of cell–cell contact, indicating the lack of a global signal transduction cascade leading to the generation of O2− over the entirety of the cells. Bar: 10 μm. Reprinted from Robinson and Badwey 1995
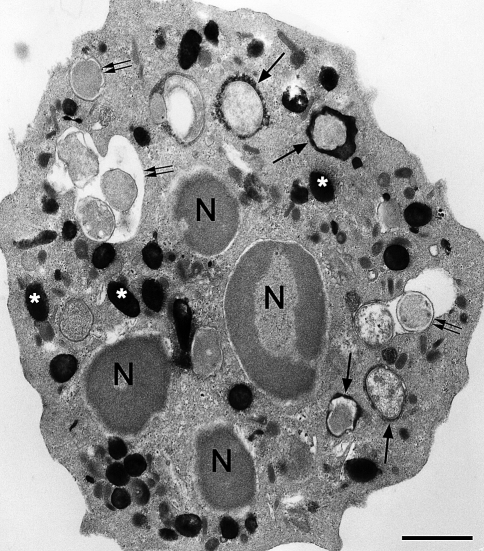
Electron micrograph of a human neutrophil that has ingested E. coli minicells; the neutrophil was subsequently reacted for the cytochemical detection of MPO activity using the DAB method with exogenous H2O2. Black electron dense deposits are present in intracellular granules, the azurophil granules (asterisks), and within a population of phagosomes (arrows). Under these incubation conditions, there were some phagosomes that had not yet acquired MPO (double arrows). The cytochemical reaction for the detection of MPO activity following ingestion of minicells was carried out as we have described (Fox et al. 1987). Bar: 1 μm
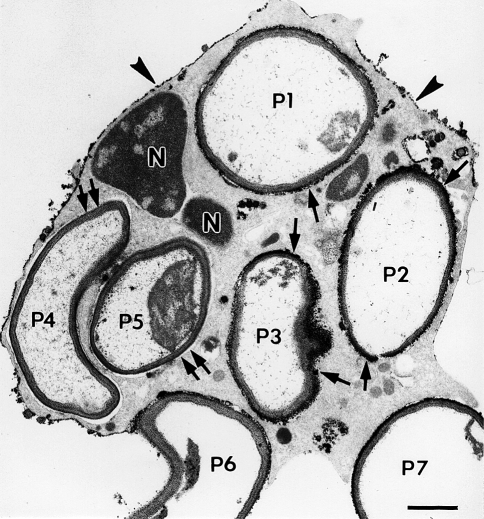
Electron micrograph illustrating the cytochemical detection of H2O2 with the cerium method in a neutrophil that was stimulated by phagocytosis. This thin-section electron micrograph is of a cell that had phagocytized zymozan particles (yeast cell walls) and was then incubated for cytochemistry as described previously (Briggs et al. 1975b). Electron-dense reaction product is present on portions of the cell surface (arrowheads) and in some phagosomes. In this cell profile, five zymozan particles appear to be within phagosomes (P1–5), while two additional particles are bound to the cell surface, but are incompletely internalized (P6–7). Note that some particles (P1–3) have reaction product surrounding the luminal aspect of the phagosomes (arrows), while other particles (P4–5) lack reaction product in the phagosomes (double arrows). Portions of the nucleus (N) are evident. These results indicate an asymmetry in the spatial and/or temporal delivery of H2O2 to phagosomes. Bar: 1 μm. Reprinted from Robinson and Badwey 1995
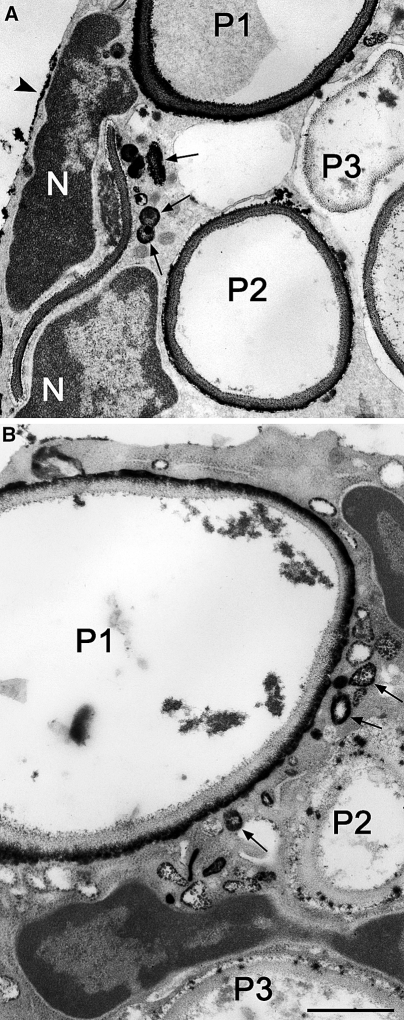
Electron micrographs of portions of two different neutrophils that ingested zymosan particles; sites of H2O2 generation were detected with the cerium-based cytochemical reaction. a A portion of a cell is shown in which phagosomes show a spectrum of responses: P1, heavy reaction product; P2, moderate reaction product; P3, little reaction product. There are several small vesicular structures containing reaction product in the periphagosome region (arrows) suggesting intracellular sites of H2O2. Patches of reaction product are at the cell surface (arrowhead). Portions of the nucleus (N) are visible. b A portion of a cell is shown in which phagosomes show different responses: P1, heavy reaction product; P2 and P3, little reaction product. There are several small vesicular structures containing reaction product in the periphagosome region (arrows) suggesting intracellular sites of H2O2. Patches of reaction product are at the cell surface (arrowhead). Portions of the nucleus (N) are visible. Bars: 1 μm
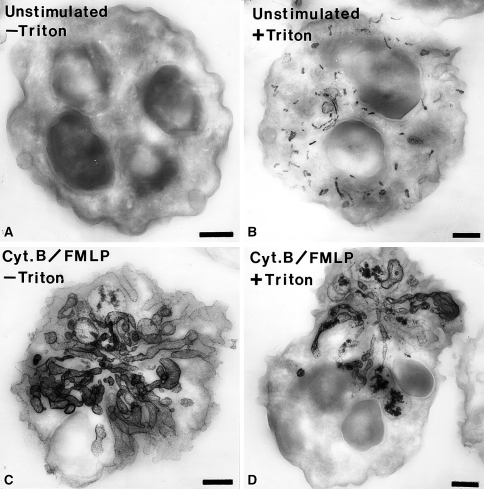
Electron micrographs of thick sections (0.35 μm thickness) of human neutrophils incubated for the cytochemical detection of alkaline phosphatase. a An unstimulated cell that was incubated for the detection of alkaline phosphatase activity in the absence of cell permeabilization with Triton X-100. Note that there is no reaction in the absence of permeabilization. b A cell that was incubated for the detection of alkaline phosphatase activity following cell permeabilization. In this case, numerous small rod-shaped organelles containing reaction product are evident. c A cell stimulated with f-Met-Leu-Phe (FMLP) and cytochalasin B (Cyt.B) that was incubated for the detection of alkaline phosphatase activity in the absence of cell-permeabilization. Note the dramatic rearrangement of the alkaline phosphatase compartments into elaborate tubular structures. These tubular structures appear to be in continuity with the extracellular space since permeabilization was not required to detect the enzyme activity. d A cell stimulated with f-Met-Leu-Phe (FMLP) and cytochalasin B (Cyt.B) that was incubated for the detection of alkaline phosphatase activity in the presence of cell permeabilization. This cell has a appearance similar to the cell treated in the same manner but without permeabilization. The cytochemical reaction was carried out as we have described previously (Kobayashi and Robinson 1991). Bars: 1 μm
Similar articles
-
Phagocytic leukocytes and reactive oxygen species.Histochem Cell Biol. 2009 Apr;131(4):465-9. doi: 10.1007/s00418-009-0565-5. Epub 2009 Feb 18. Histochem Cell Biol. 2009. PMID: 19224236 Review.
-
Measurement of Respiratory Burst Products, Released or Retained, During Activation of Professional Phagocytes.Methods Mol Biol. 2020;2087:301-324. doi: 10.1007/978-1-0716-0154-9_22. Methods Mol Biol. 2020. PMID: 31729000
-
Cell-Free NADPH Oxidase Activation Assays: A Triumph of Reductionism.Methods Mol Biol. 2020;2087:325-411. doi: 10.1007/978-1-0716-0154-9_23. Methods Mol Biol. 2020. PMID: 31729001 Review.
-
NADPH oxidase-mediated generation of reactive oxygen species is critically required for survival of undifferentiated human promyelocytic leukemia cell line HL-60.Free Radic Res. 2004 Jun;38(6):629-37. doi: 10.1080/10715760410001694053. Free Radic Res. 2004. PMID: 15346654
-
Chronic Granulomatous Disease.Methods Mol Biol. 2019;1982:531-542. doi: 10.1007/978-1-4939-9424-3_32. Methods Mol Biol. 2019. PMID: 31172494 Review.
Cited by
-
Effect of Byrsonima crassa and phenolic constituents on Helicobacter pylori-induced neutrophils oxidative burst.Int J Mol Sci. 2012;13(1):133-41. doi: 10.3390/ijms13010133. Epub 2011 Dec 23. Int J Mol Sci. 2012. PMID: 22312243 Free PMC article.
-
Oxidative stress induces inflammation of lens cells and triggers immune surveillance of ocular tissues.Chem Biol Interact. 2022 Mar 1;355:109804. doi: 10.1016/j.cbi.2022.109804. Epub 2022 Feb 4. Chem Biol Interact. 2022. PMID: 35123994 Free PMC article.
-
Genotoxic Potential of Nanoparticles: Structural and Functional Modifications in DNA.Front Genet. 2021 Sep 29;12:728250. doi: 10.3389/fgene.2021.728250. eCollection 2021. Front Genet. 2021. PMID: 34659351 Free PMC article. Review.
-
Neutrophils: Beneficial and Harmful Cells in Septic Arthritis.Int J Mol Sci. 2018 Feb 5;19(2):468. doi: 10.3390/ijms19020468. Int J Mol Sci. 2018. PMID: 29401737 Free PMC article. Review.
-
State-of-the-art technologies, current opinions and developments, and novel findings: news from the field of histochemistry and cell biology.Histochem Cell Biol. 2008 Dec;130(6):1205-51. doi: 10.1007/s00418-008-0535-3. Epub 2008 Nov 5. Histochem Cell Biol. 2008. PMID: 18985372 Review.
References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PMC', 'value': 'PMC1137899', 'is_inner': False, 'url': 'https://pmc.ncbi.nlm.nih.gov/articles/PMC1137899/'}, {'type': 'PubMed', 'value': '8141770', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/8141770/'}]}
- Abo A, Webb MR, Grogan A, Segal AW (1994) Activation of NADPH oxidase involves the dissociation of p21rac from its inhibitory GDP/GTP exchange protein (rhoGDI) followed by its translocation to the plasma membrane. Biochem J 298:585–591 - PMC - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '10233905', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/10233905/'}]}
- Allen L-AH, DeLeo FR, Gallois A, Toyoshima S, Suzuki K, Nauseef WM (1999) Transient association of the nicotinamide adenine dinucleotide phosphate oxidase subunits p47phox and p67phox with phagosomes in neutrophils from patients with X-linked chronic granulomatous disease. Blood 93:3521–3530 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '1259763', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/1259763/'}]}
- Allen RC, Loose LD (1976) Phagocytic activation of a luminol-dependent chemiluminescence in rabbit alveolar and peritoneal macrophages. Biochem Biophys Res Commun 69:245–252 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '5026288', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/5026288/'}]}
- Allen RC, Stjernholm RL, Steele RH (1972) Evidence for the generation of an electronic excitation state(s) in human polymorphonuclear leukocytes and its participation in bactericidal activity. Biochem Biophys Res Commun 47:679–684 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'PubMed', 'value': '15059619', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/15059619/'}]}
- Ambruso DR, Cusack N, Thurman G (2004) NADPH oxidase activity of neutrophil specific granules: requirements for cytodolic components and evidence of assembly during cell activation. Mol Genet Metab 81:313–321 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
