

The Poly(ADP-ribose) polymerase PARP-1 is required for oxidative stress-induced TRPM2 activation in lymphocytes
- PMID: 18599483
- PMCID: PMC3259813
- DOI: 10.1074/jbc.M802673200
The Poly(ADP-ribose) polymerase PARP-1 is required for oxidative stress-induced TRPM2 activation in lymphocytes
Abstract
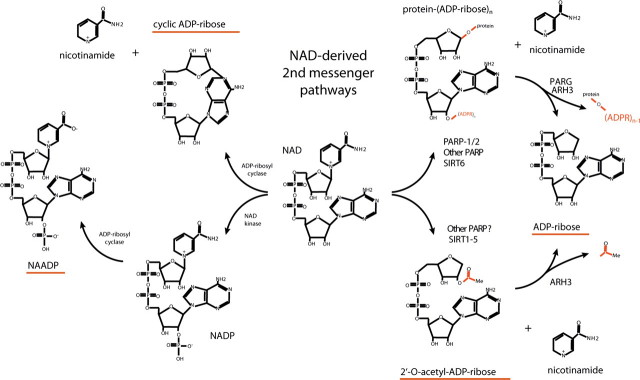
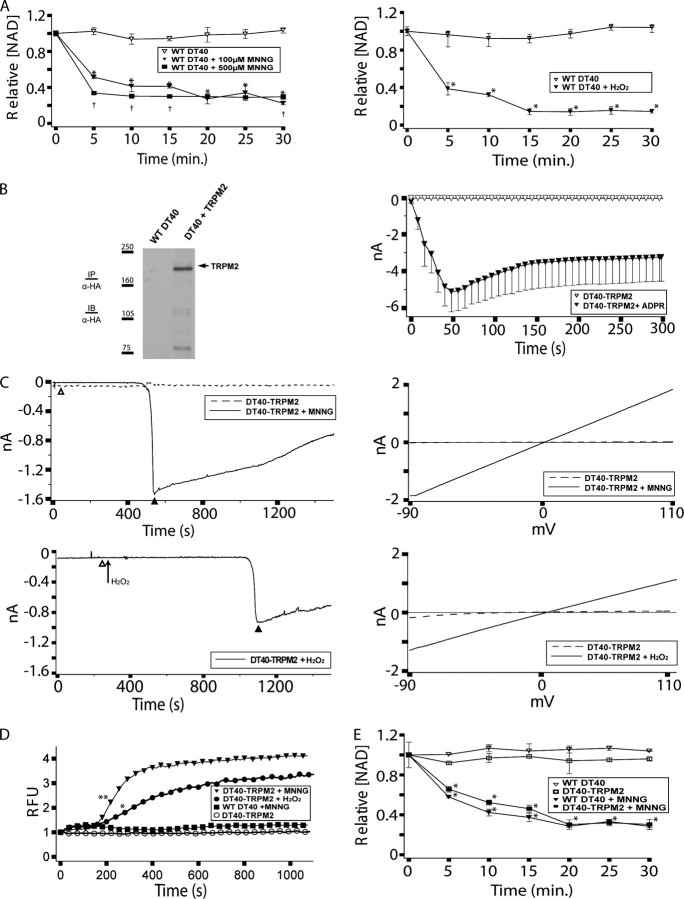
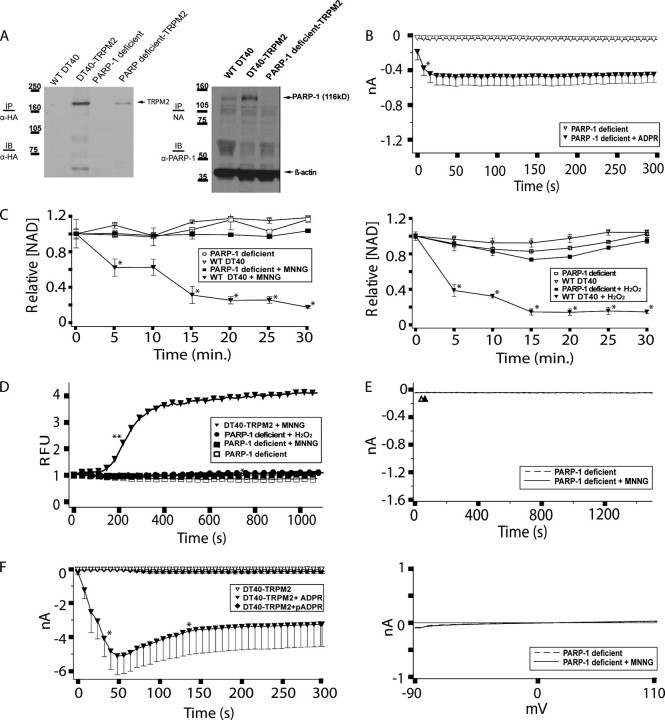
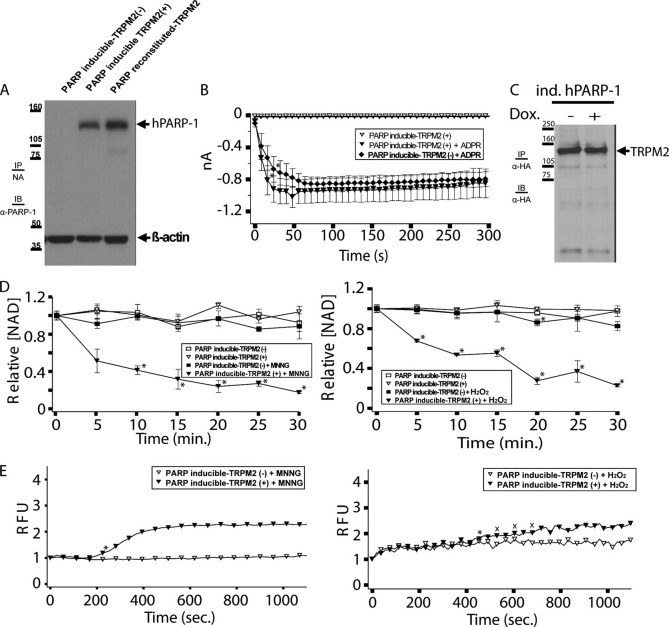
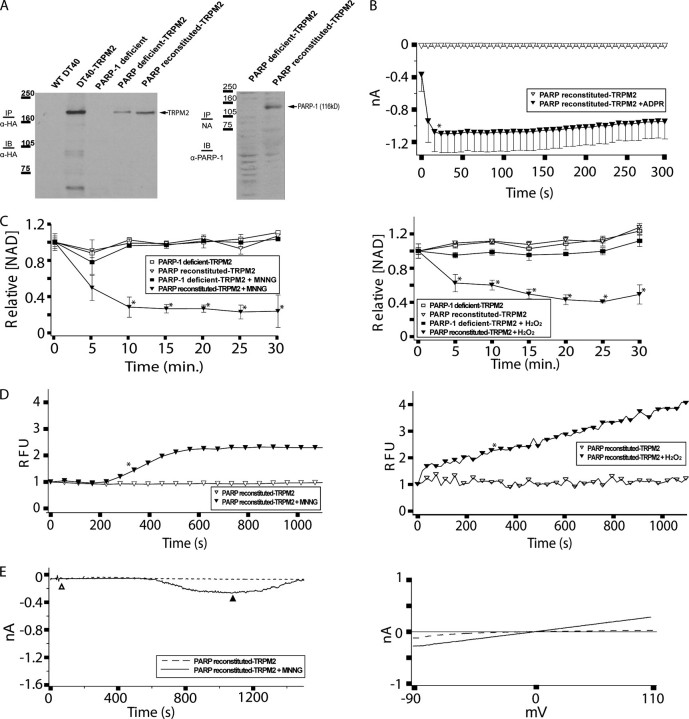
TRPM2 cation channels are widely expressed in the immune system and are thought to play a role in immune cell responses to oxidative stress. Patch clamp analyses suggest that TRPM2 channel activation can occur through a direct action of oxidants on TRPM2 channels or indirectly through the actions of a related group of adenine nucleotide 2nd messengers. However, the contribution of each gating mechanism to oxidative stress-induced TRPM2 activation in lymphocytes remains undefined. To better understand the molecular events leading to TRPM2 activation in lymphocytes, we analyzed oxidative stress-induced turnover of intracellular NAD, the metabolic precursor of adenine nucleotide 2nd messengers implicated in TRPM2 gating, and oxidative stress-induced TRPM2-mediated currents and Ca2+ transients in DT40 B cells. TRPM2-dependent Ca2+ entry did not influence the extent or time course of oxidative stress-induced turnover of NAD. Furthermore, expression of oxidative stress-activated poly(ADP-ribose) polymerases (PARPs) was required for oxidative stress-induced NAD turnover, TRPM2 currents, and TRPM2-dependent Ca2+ transients; no oxidant-induced activation of TRPM2 channels could be detected in PARP-deficient cells. Together, our results suggest that during conditions of oxidative stress in lymphocytes, TRPM2 acts as a downstream effector of the PARP/poly(ADP-ribose) glycohydrolase pathway through PARP-dependent formation of ADP-ribose.
Figures
References
-
- Inada, H., Iida, T., and Tominaga, M. (2006) Biochem. Biophys. Res. Commun. 350 762-767 - PubMed
-
- Gasser, A., Bruhn, S., and Guse, A. H. (2006) J. Biol. Chem. 281 16906-16913 - PubMed
-
- Beck, A., Kolisek, M., Bagley, L. A., Fleig, A., and Penner, R. (2006) FASEB J. 20 962-964 - PubMed
-
- Gasser, A., Glassmeier, G., Fliegert, R., Langhorst, M. F., Meinke, S., Hein, D., Kruger, S., Weber, K., Heiner, I., Oppenheimer, N., Schwarz, J. R., and Guse, A. H. (2006) J. Biol. Chem. 281 2489-2496 - PubMed
-
- Kraft, R., Grimm, C., Grosse, K., Hoffmann, A., Sauerbruch, S., Kettenmann, H., Schultz, G., and Harteneck, C. (2004) Am. J. Physiol. 286 C129-C137 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
