

Pontine respiratory-modulated activity before and after vagotomy in decerebrate cats
- PMID: 18599543
- PMCID: PMC2652175
- DOI: 10.1113/jphysiol.2008.152108
Pontine respiratory-modulated activity before and after vagotomy in decerebrate cats
Erratum in
- J Physiol. 2008 Nov 1;586(21):5279
Abstract
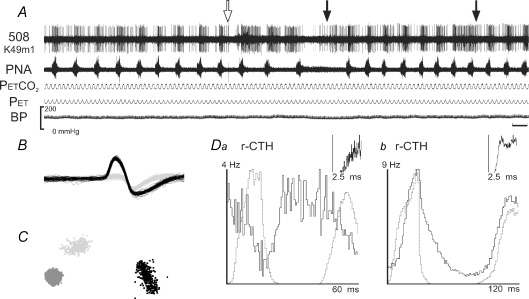
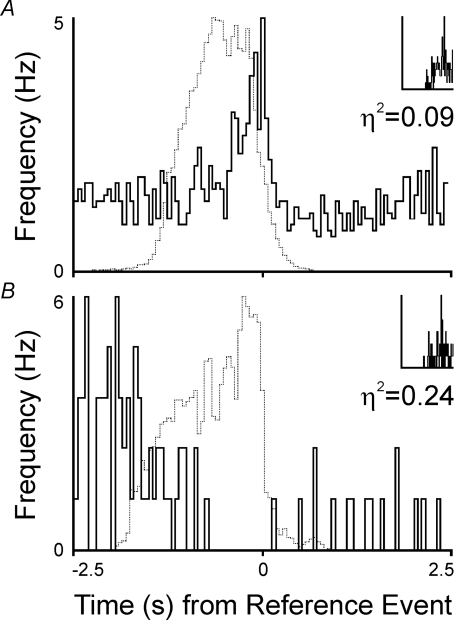
The dorsolateral (DL) pons modulates the respiratory pattern. With the prevention of lung inflation during central inspiratory phase (no-inflation (no-I or delayed-I) tests), DL pontine neuronal activity increased the strength and consistency of its respiratory modulation, properties measured statistically by the eta(2) value. This increase could result from enhanced respiratory-modulated drive arising from the medulla normally gated by vagal activity. We hypothesized that DL pontine activity during delayed-I tests would be comparable to that following vagotomy. Ensemble recordings of neuronal activity were obtained before and after vagotomy and during delayed-I tests in decerebrate, paralysed and ventilated cats. In general, changes in activity pattern during the delayed-I tests were similar to those after vagotomy, with the exception of firing-rate differences at the inspiratory-expiratory phase transition. Even activity that was respiratory-modulated with the vagi intact became more modulated while withholding lung inflation and following vagotomy. Furthermore, we recorded activity that was excited by lung inflation as well as changes that persisted past the stimulus cycle. Computer simulations of a recurrent inhibitory neural network model account not only for enhanced respiratory modulation with vagotomy but also the varied activities observed with the vagi intact. We conclude that (a) DL pontine neurones receive both vagal-dependent excitatory inputs and central respiratory drive; (b) even though changes in pontine activity are transient, they can persist after no-I tests whether or not changes in the respiratory pattern occur in the subsequent cycles; and (c) models of respiratory control should depict a recurrent inhibitory circuitry, which can act to maintain the stability and provide plasticity to the respiratory pattern.
Figures









References
-
- Alheid GF, Milsom WK, McCrimmon DR. Pontine influences on breathing: an overview. Respir Physiol Neurobiol. 2004;143:105–114. - PubMed
-
- Baekey DM, Morris KF, Nuding S, Segers LS, Lindsey BG, Shannon R. Ventrolateral medullary respiratory network participation in the expiration reflex in the cat. J Appl Physiol. 2004;96:2057–2072. - PubMed
-
- Balis UJ, Morris KF, Koleski J, Lindsey BG. Simulations of a ventrolateral medullary neural network for respiratory rhythmogenesis inferred from spike train cross-correlation. Biol Cybern. 1994;70:311–327. - PubMed
-
- Berman AL. The Brain Stem of the Cat: A Cytoarchitectonic Atlas with Stereotaxic Coordinates. Madison: University of Wisconsin Press; 1968.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous