

Review
doi: 10.1016/j.sbi.2008.05.008.
Epub 2008 Jul 23.
Ligand binding by repeat proteins: natural and designed
Affiliations
- PMID: 18602006
- PMCID: PMC3500881
- DOI: 10.1016/j.sbi.2008.05.008
Item in Clipboard
Review
Ligand binding by repeat proteins: natural and designed
Curr Opin Struct Biol.
2008 Aug.
Abstract
Repeat proteins contain tandem arrays of small structural motifs. As a consequence of this architecture, they adopt non-globular, extended structures that present large, highly specific surfaces for ligand binding. Here we discuss recent advances toward understanding the functional role of this unique modular architecture. We showcase specific examples of natural repeat proteins interacting with diverse ligands and also present examples of designed repeat protein-ligand interactions.
Figures
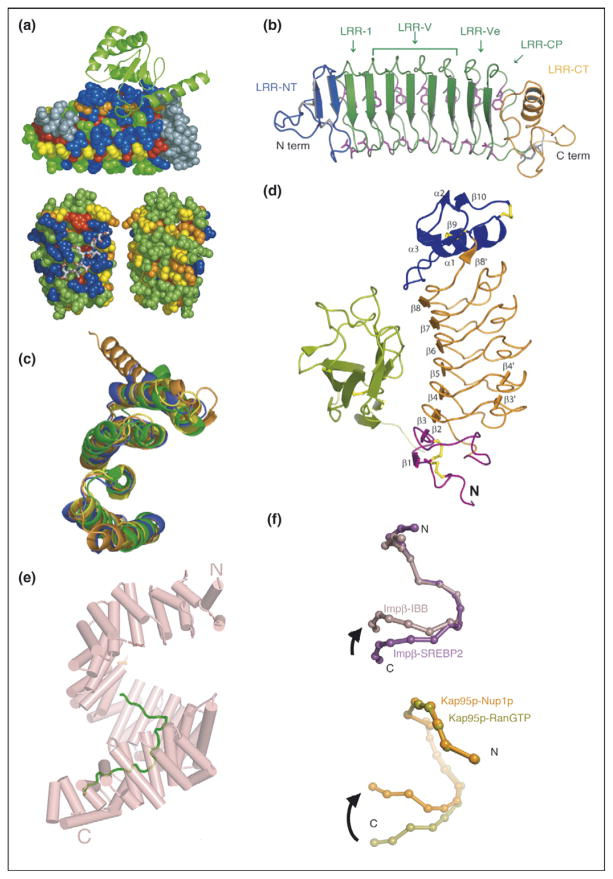
Repeat proteins as conserved scaffolds. (a) Sequence variability analysis repeat proteins. Upper panel: variability analysis of 15 497 Ank repeats. The 4-Ank domain from GABPβ1 (spheres) is shown bound to the ligand GABPα (ribbons) (PBD ID: 1awc). Coloring is from most variable = blue to least variable = red. The binding surface is clearly more blue than the rest of the proteins’ surface. Lower panel: sequence variability analysis of 6887 TPR repeats mapped onto the co-crystal structures of HOP-TPR1/Hsc70 peptide (PBD ID: 1elw) with the TPR domain rendered in spheres and the ligands in sticks. Two views from 180° rotation of the molecule are shown. The concave, ligand binding surface, left, is clearly more variable (blue) than the convex, solvent exposed surfaces (green, yellow), right [1,3]. (b) Crystal structure of hagfish variable lymphocyte receptor (VLR-A). The VLR-A protein comprises 8 LLR modules. The constant N-terminal (LRR-NT) and C-terminal (LRR-CT) modules are shown in blue and orange, respectively. The variable LRR modules (LRR-V) that can be present in variable numbers are colored in green [2••]. (c) Alignment of TPR domains. Structural alignment of TPR domains unliganded and peptide-bound: CTPR3 (PDB ID 1NA0) [35] in blue, TPR domain (residues 19-177) of PP5 in orange (PDB ID: 1a17) [36], and TPR2A (PDB ID: 1elr) in green and TPR1 (PDB ID: 1elw) in yellow, domains of Hop from the co-crystal structures with their peptide ligands, C-terminal peptides of Hsp90 and Hsp70, respectively [37]. The structures are all completely superimposable, with backbone RMSD values that vary from 1.1 to 1.9 Å for different pairwise structural alignments [5]. (d) Structure of Slit2 D2 bound to Robo1 Ig1. Ig1 is in green; Slit2 D2 N-terminal and C-terminal caps are in purple and blue, respectively; LRRs 1–6 are in orange; and the disulfide bridges are in yellow [6]. (e) Structure of HEAT domain of karyopherin Kapβ2 bound to NLS substrate (PDB ID: 1ot8). The helices are represented as cylinders and structurally disordered loops as dashed lines. The extended NLS sequence of the cargo protein hnRNPA1 is shown in green. A1-NLS peptide binds in an extended conformation on the concave surface of the C-terminal arch of the karyopherin superhelix [10••]. (f) Schematic illustration of the hinge motion between subgroups of HEAT repeats. (Top) Schematic illustration of the hinge movement between HEAT repeat subdomains H1–H12 and H14–H19 in Impβ and Kap95p upon ligand binding. IMM-bound Impβ (pink) and SREBP2-bound Impβ (purple) are superimposed at HEAT repeats 5–10, with a sphere representing the center of each HEAT repeat. (Bottom) Schematic illustration of the hinge movement between HEAT repeat subdomains H1–H14 and H15–H19. RanGTP-bound Kap95p (light green) and Nup1p-bound Kap95p (orange) are superimposed at HEAT repeats 6–13, with a sphere representing the center of each HEAT repeat [10••].
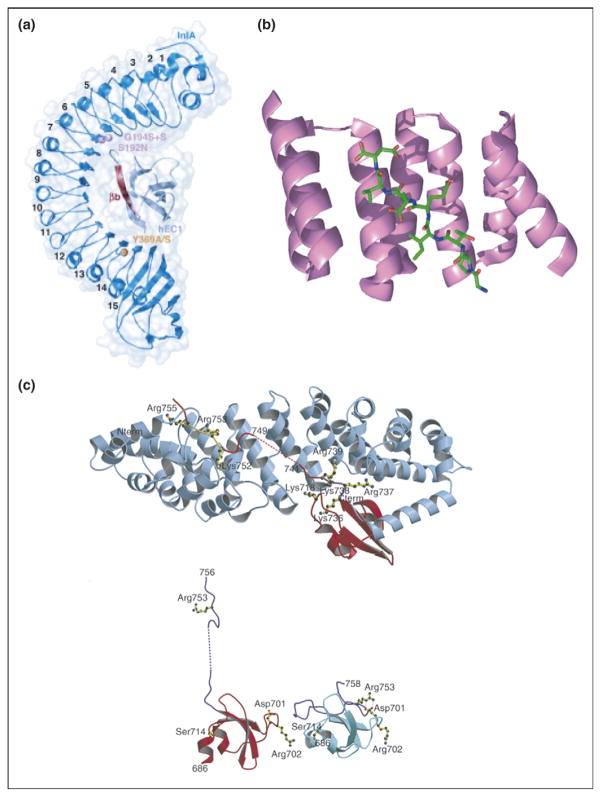
Repeat proteins bind to extended ligands. (a) Overall structure of internalin (InlA)/N-terminal domain of human E-cadherin (hEC1) complex shown to be in dark and light blue, respectively. The eight LRRs of InlA interact with a long, extended, β-strand of hEC1 [13••]. (b) Crystal structure of TPR1 domain of HOP in complex with the extended C-terminal peptide of Hsp70 bound in the concave face of the TPR domain. TPR1 domain is shown as a ribbon (purple) and the 8-mer C-terminal peptide of Hsp70 is shown as sticks (green) [37]. (c) (Top) Ribbon diagram showing C-terminal domain of influenza virus polymerase PB2 subunit (red) bound to human importin α5 (blue), comprising 10 armadillo repeats. (Bottom) Comparison of the PB2 domain structure in complexed (red) and free solution state (cyan) demonstrates unfolding of residues 736–759 (purple) upon binding to importin α5 [14••].
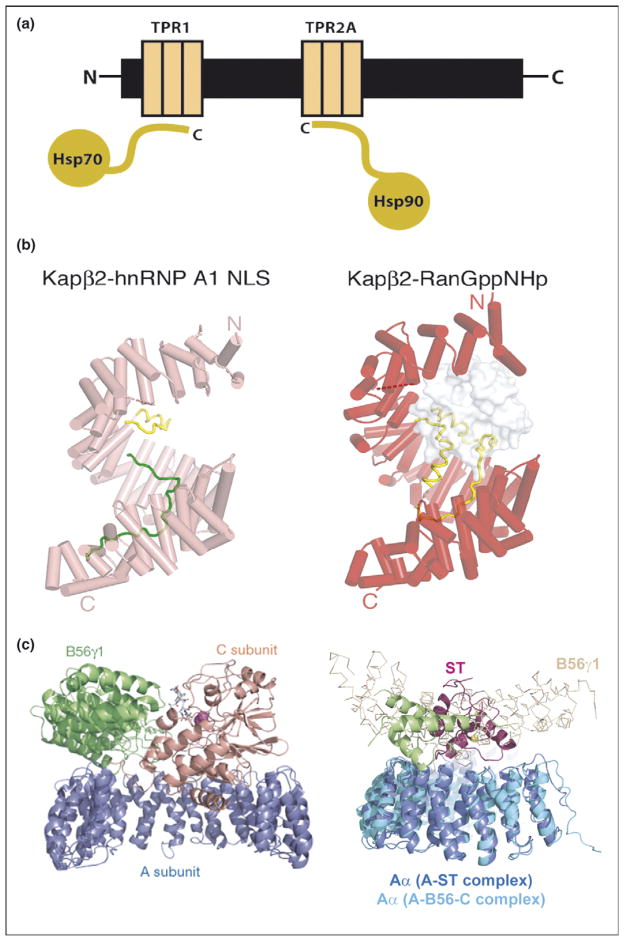
Repeat proteins as multi-protein complex organizers. (a) Schematic representation of Hsp90/Hsp70 organizing protein (HOP) showing how two independent TPR domains serve to assemble the multi-chaperone complex. The figure shows the interactions between the TPR1 and TPR2A domain of HOP with the C-terminal sequences of Hsp70 and Hsp90, respectively. (b) Heat repeats of karyopherins present different binding surfaces for multi-protein binding. Structure of karyopherin Kapβ2 bound to NLS substrate (PDB ID: 1ot8) (left panel) and bound to the regulatory protein Ran (PDB ID: 1qbk) (right panel). The helices are represented as cylinders (pink in susbtrate-bound structure and red in the Ran-bound structure); structurally disordered loops are shown as dashed lines and HEAT repeat 8 (H8) loops are in yellow. The substrate hnRNP A1-NLS is shown as a green ribbon and the Ran protein as a surface representation in gray [10••]. (c) Protein phosphatase 2A (PP2A) HEAT repeats as scaffolding subunit. Left panel: Ribbon representation of the heterotrimeric PP2A holoenzyme crystal structure. The scaffolding subunit A colored in blue binds the regulatory subunit B56γ1 (green) and the catalytic subunit C (orange). Right panel: SV 40 Small T antigen competes with the regulatory subunit B. Structural superimposition of the A-ST complex (PDB ID: 2pf4) and the A-B56-C PP2A holoenzyme complex (PDB ID: 2iae). The two complexes are superimposed using A subunit HEAT repeats 2–10. The J and unique domain of ST are colored green and pink, respectively. The Cα trace of B56γ1 regulatory subunit are shown in light orange. It is clear that ST and B56γ1 bind to the same sites on PP2A A subunit [16••,17••].
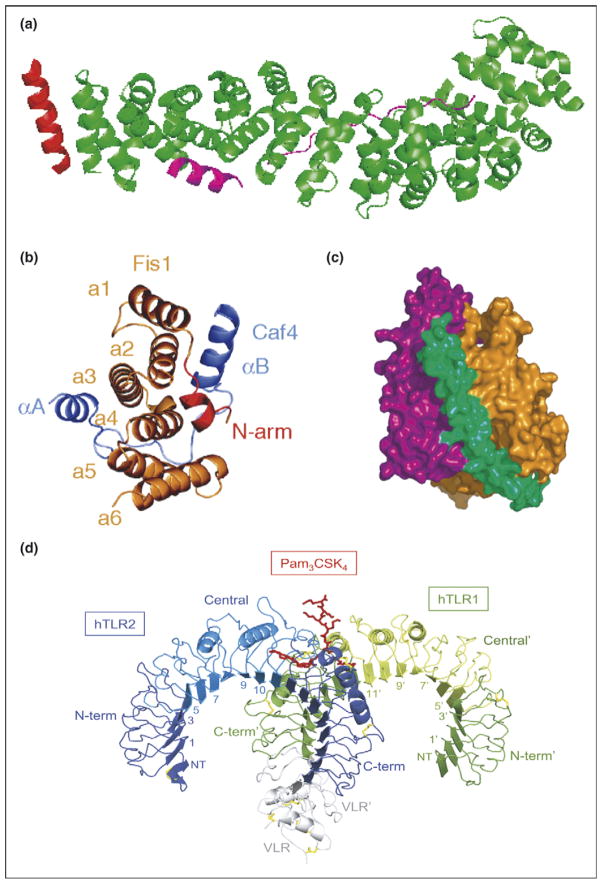
Typical and atypical modes of binding. (a) Overall structure of the β-catenin/BCL9/Tcf-4 complex in green, magenta, and red, respectively. Packing of BCL9 against the groove formed between H2 and H3 of the N-terminal capping armadillo repeat to form a helical bundle illustrates atypical binding interface for armadillo repeat proteins [8]. (b) Fis1 TPR-Caf4 complex show a new two surfaces binding mode. Crystal structures of the TPR-like protein Fis1 in complex with Caf4 protein (PDB ID: 2pqr). The Fis1 N-terminal arm (red) remains packed against the original hydrophobic groove and stabilizes the B helix of Caf4 (purple), which packs against a second hydrophobic groove on the concave surface of Fis1. In addition to the B helix packed against the concave Fis1 surface, Caf4 (purple) uses the A helix and the intervening loop to bind the convex Fis1 surface [18]. (c) Molecular surface representation of CSL–Notch–Mastermind ternary complex. CSL (orange) and Notch (purple) interact to present composite surface for binding of Mastermind (green). (d) Overall structure of the human TLR1–TLR2–Pam3CSK4 complex. The TLR1 fragments and the TLR2 fragments are shown schematically in green and blue, respectively. The central domains are colored in light green or light blue, and the Pam3CSK4 lipopeptide in red. Disulfide bridges are represented as yellow lines. Domains belonging to the TLR1 hybrid proteins are labeled with apostrophes [21••].
References
-
- Magliery TJ, Regan L. Beyond consensus: statistical free energies reveal hidden interactions in the design of a TPR motif. J Mol Biol. 2004;343:731–745. - PubMed
-
- Kim HM, Oh SC, Lim KJ, Kasamatsu J, Heo JY, Park BS, Lee H, Yoo OJ, Kasahara M, Lee JO. Structural diversity of the hagfish variable lymphocyte receptors. J Biol Chem. 2007;282:6726–6732. Inserting multiple LRR repeats from a germline repertoire to accomplish diversity. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
