

Cetacean sleep: an unusual form of mammalian sleep
- PMID: 18602158
- PMCID: PMC8742503
- DOI: 10.1016/j.neubiorev.2008.05.023
Cetacean sleep: an unusual form of mammalian sleep
Abstract
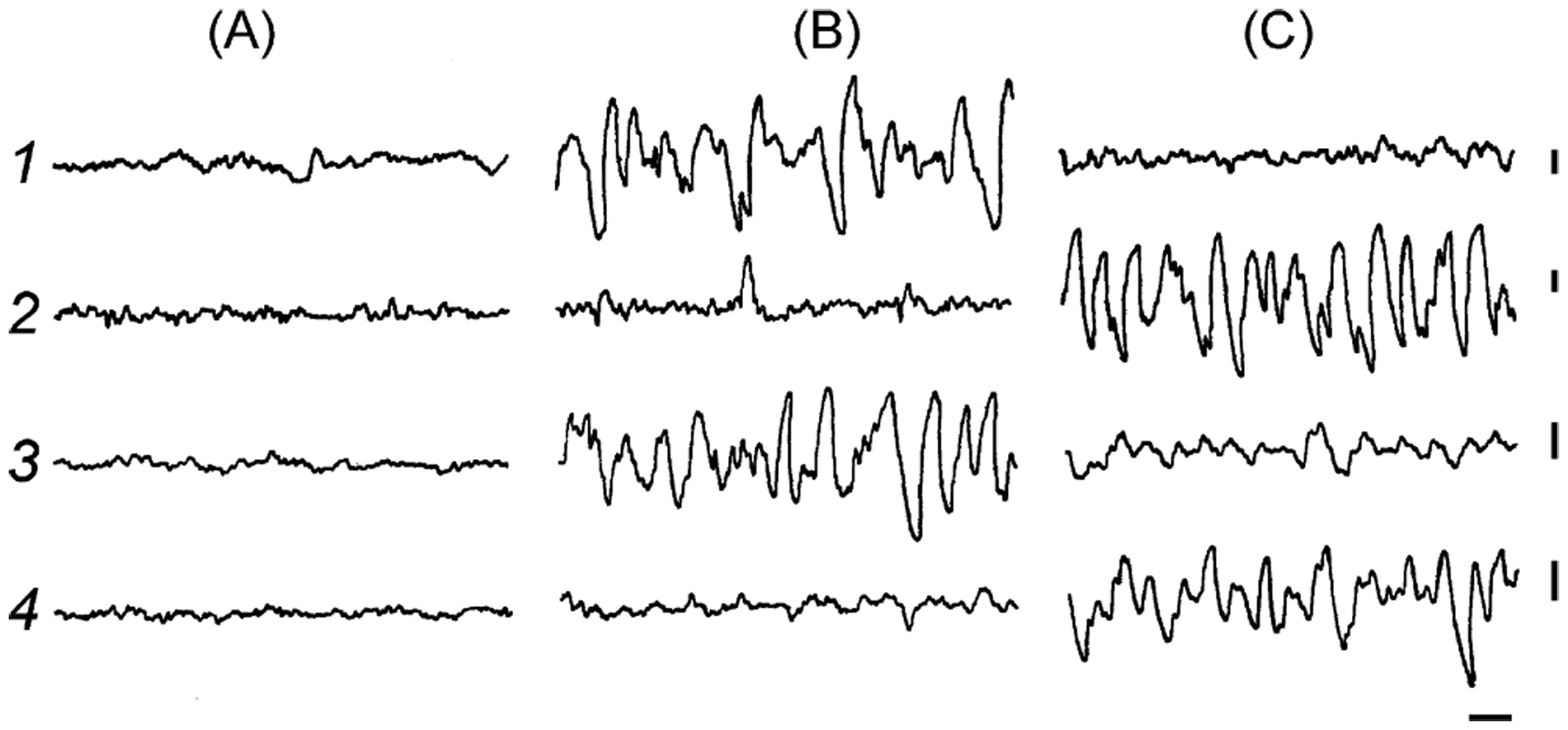
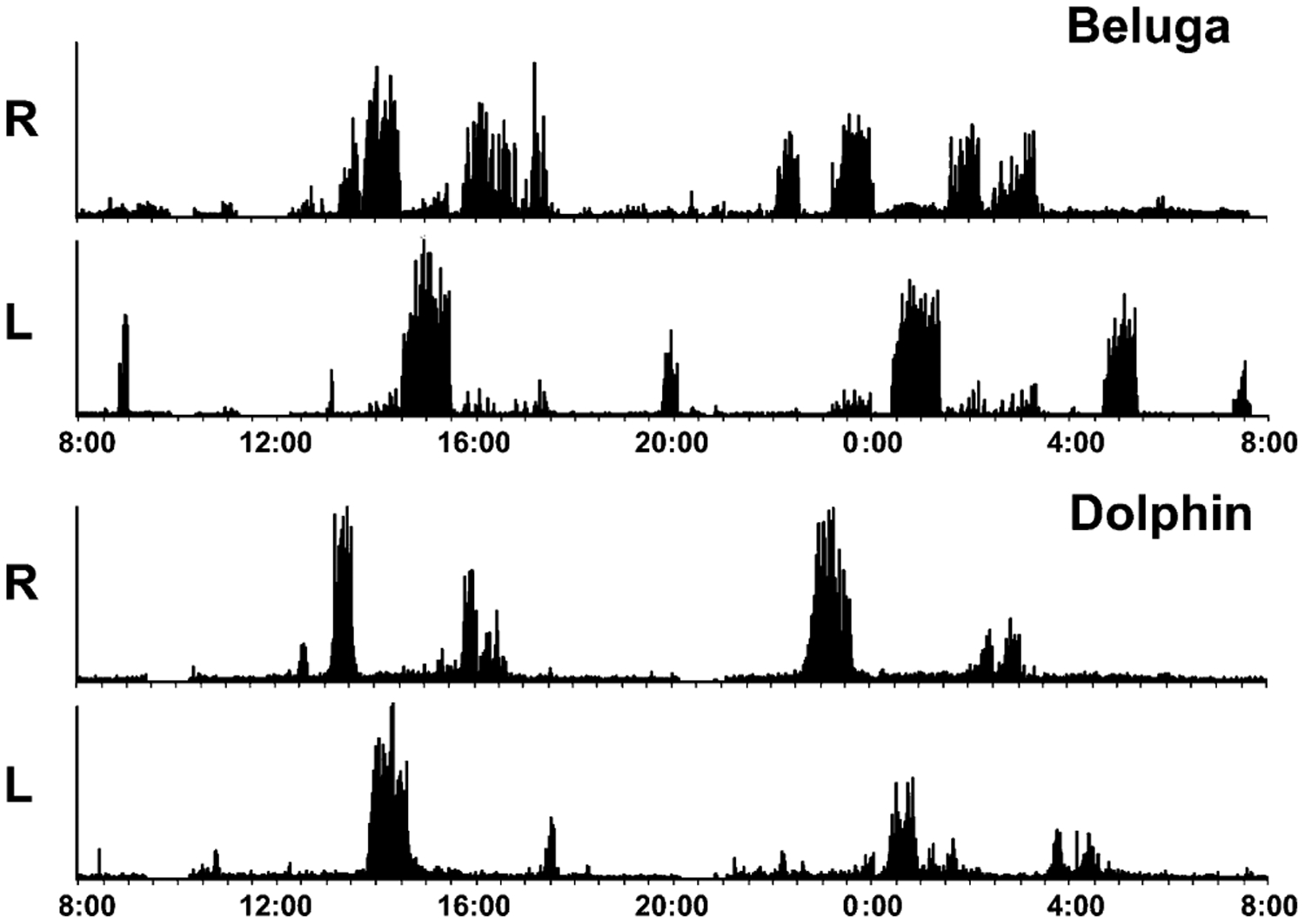
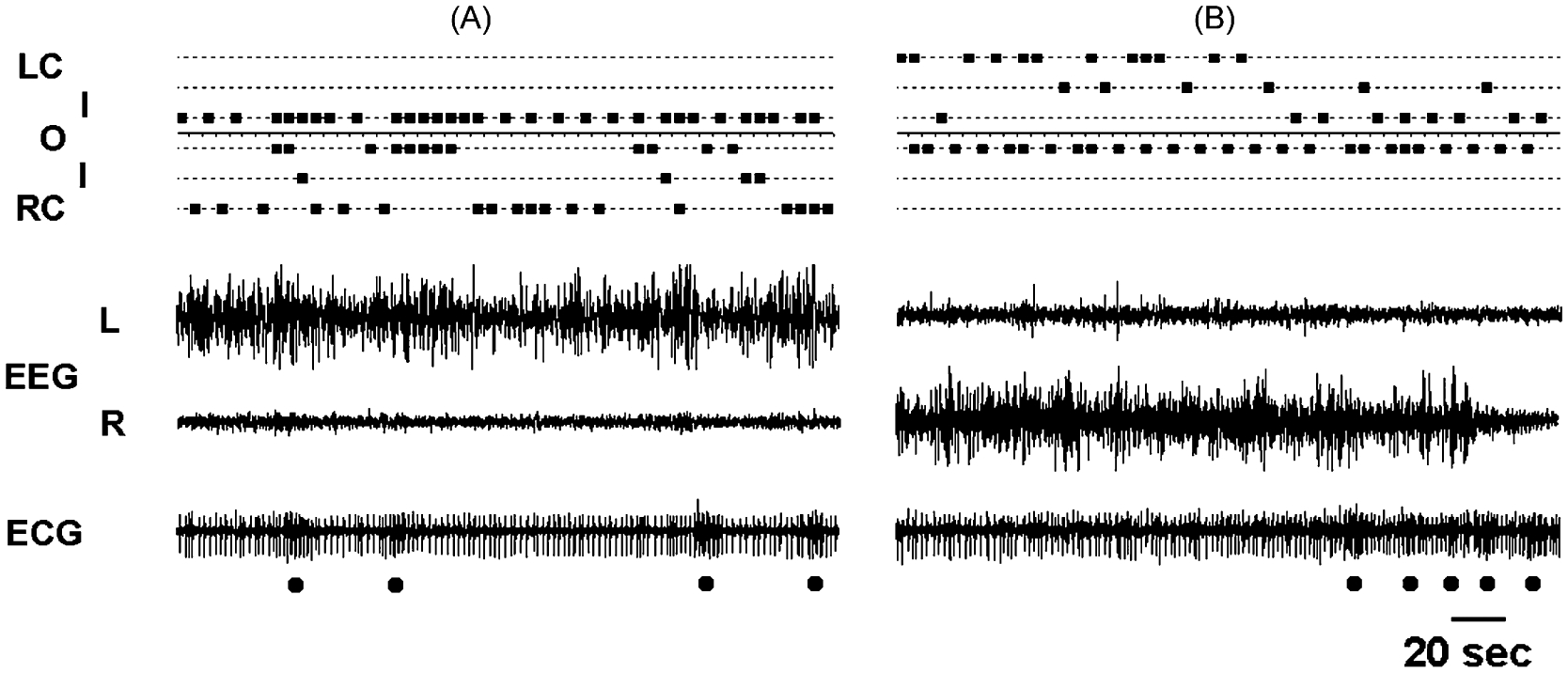
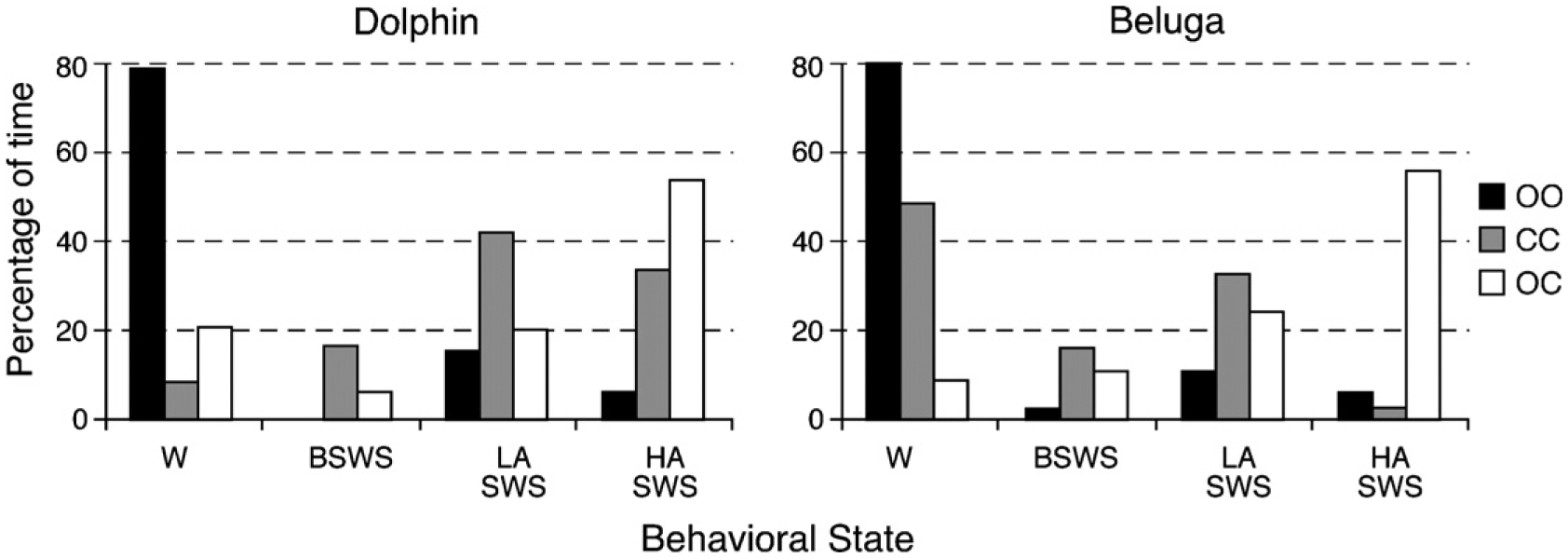
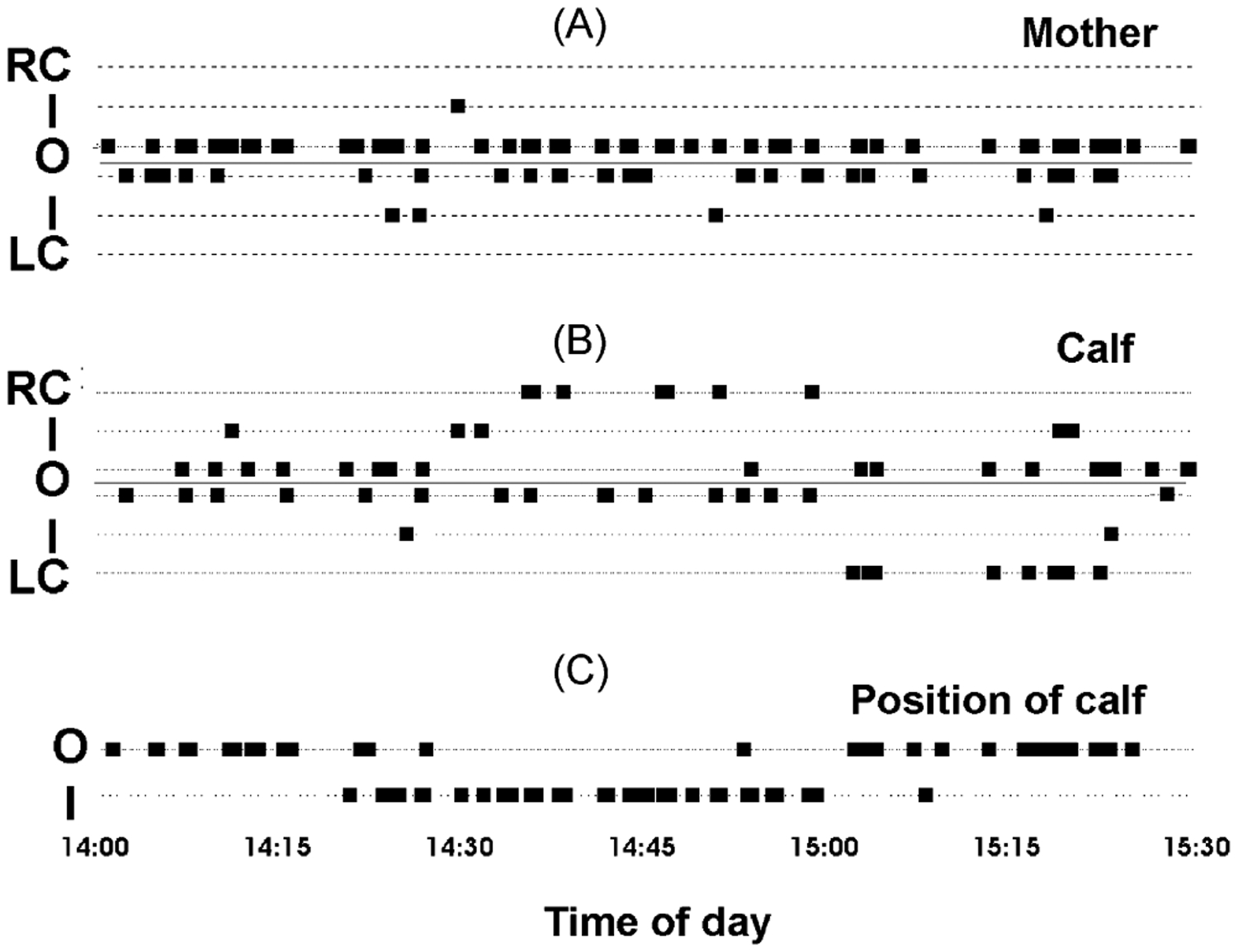
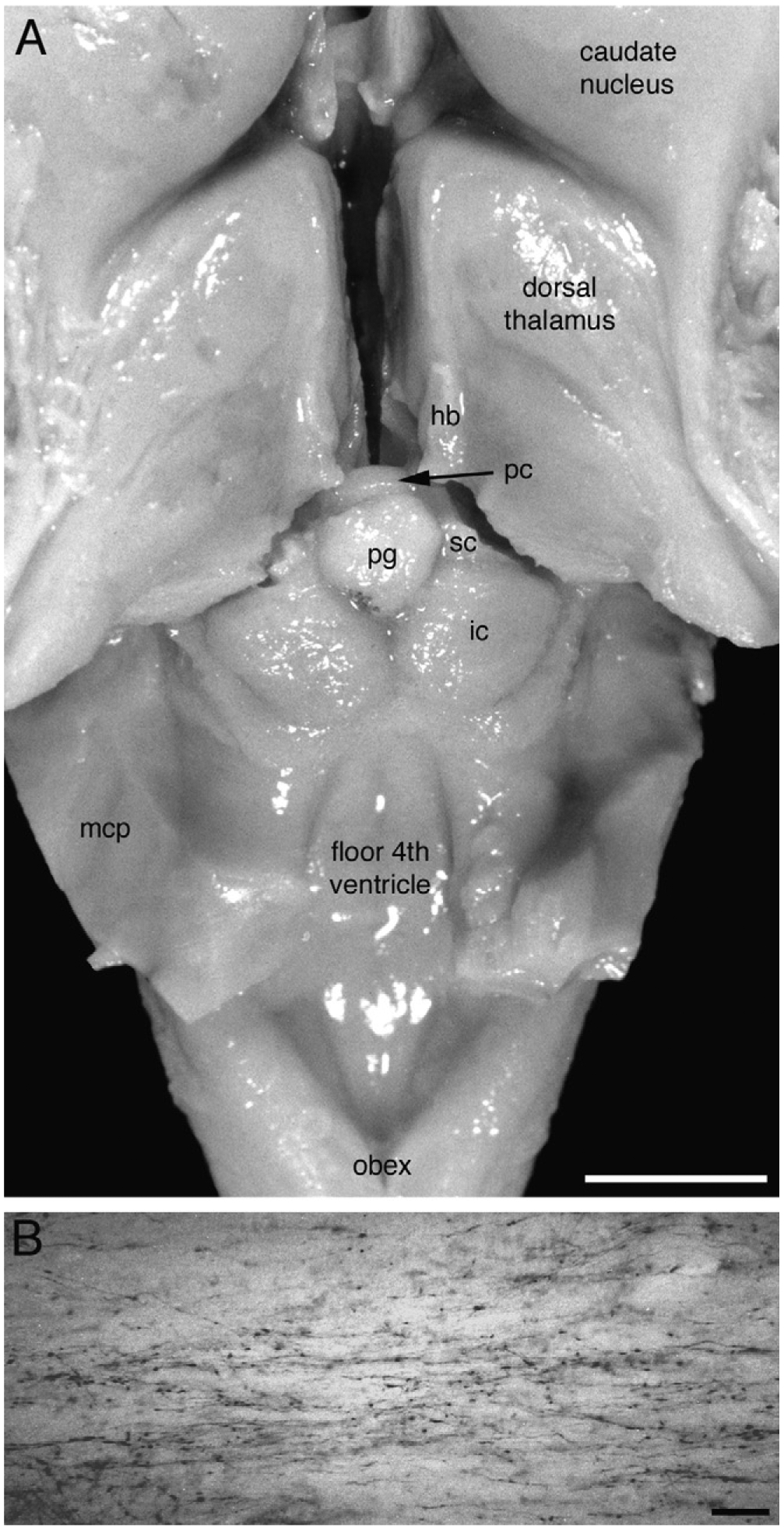
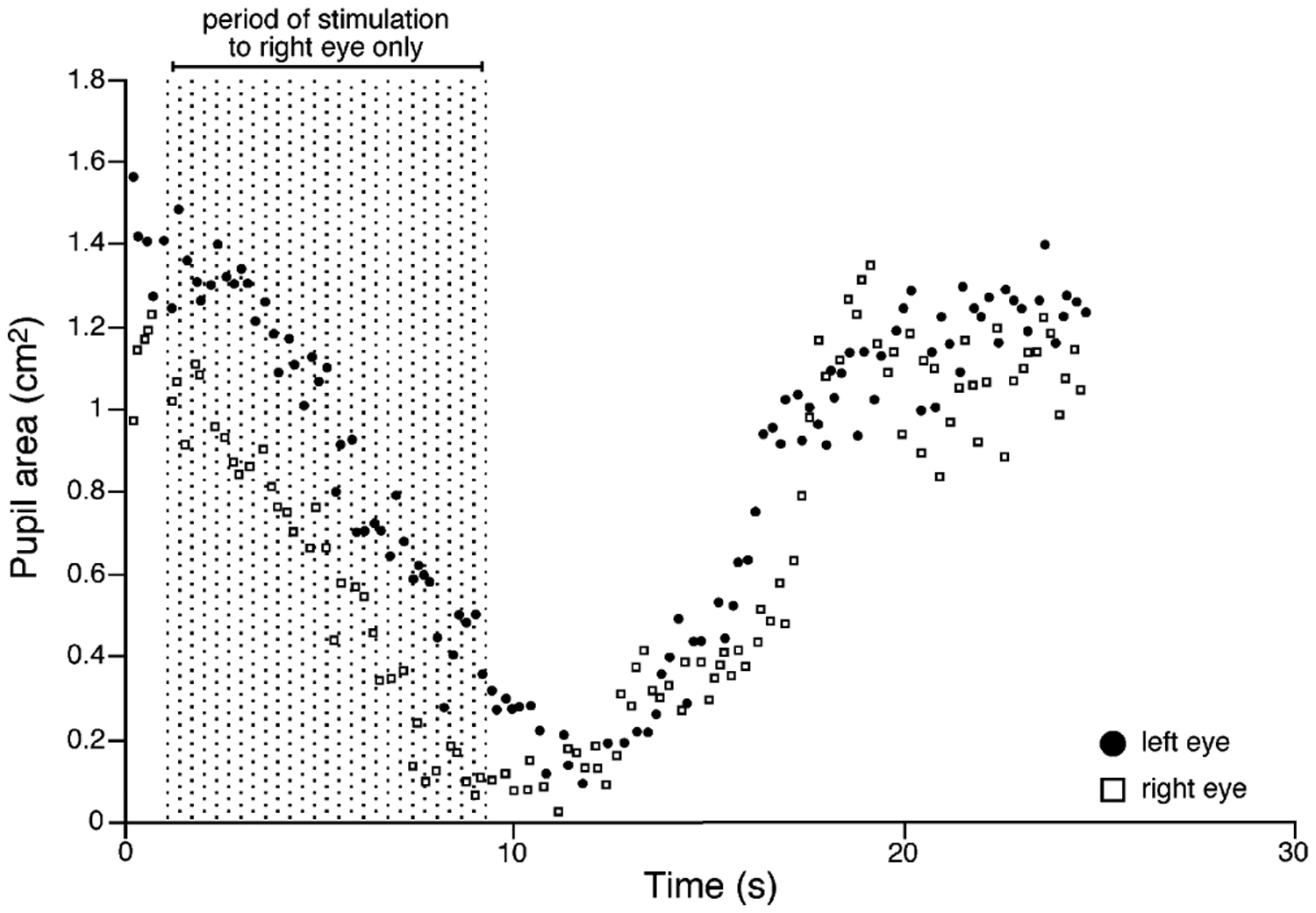
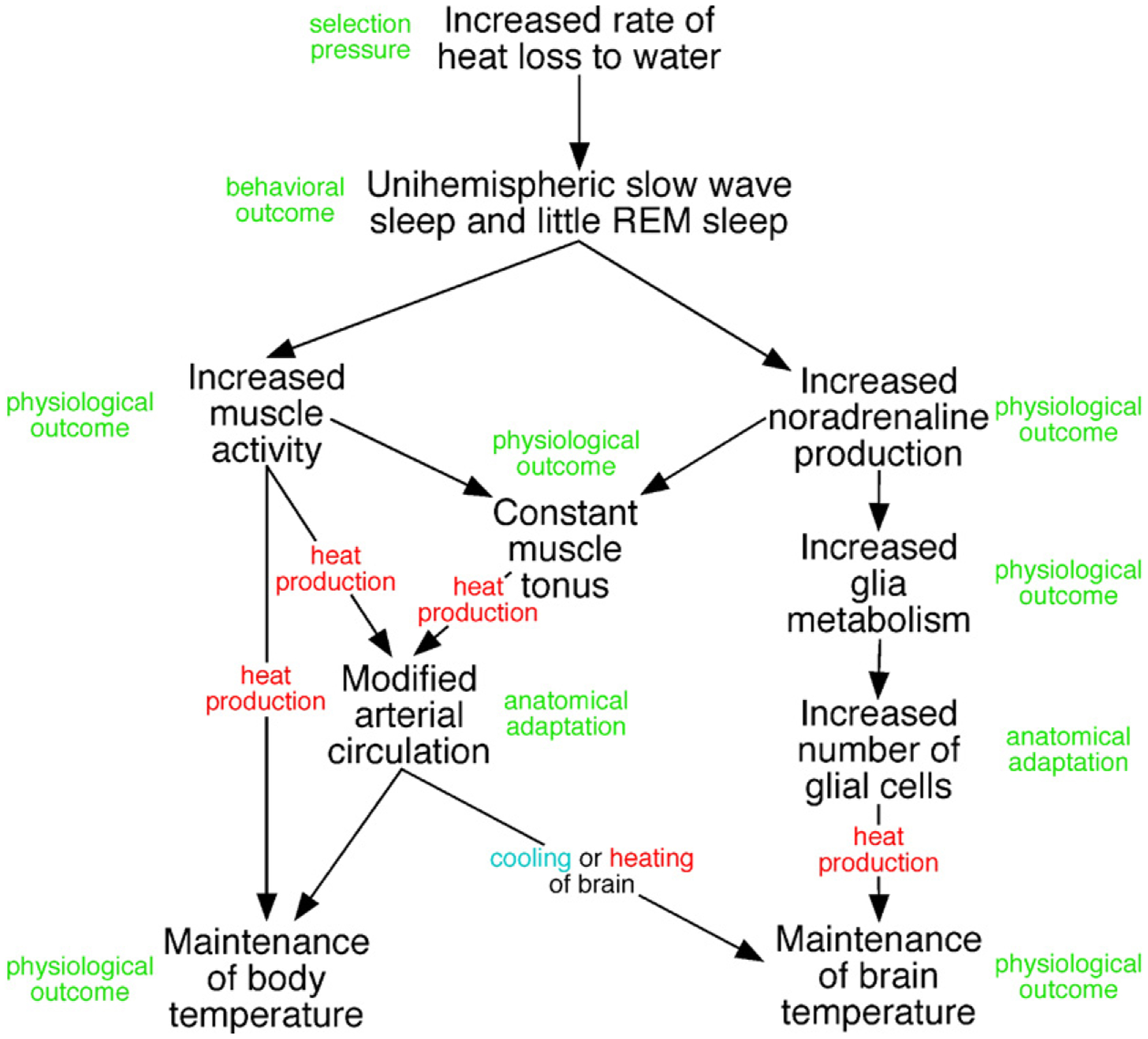
Our knowledge of the form of lateralized sleep behavior, known as unihemispheric slow wave sleep (USWS), seen in all members of the order Cetacea examined to date, is described. We trace the discovery of this phenotypically unusual form of mammalian sleep and highlight specific aspects that are different from sleep in terrestrial mammals. We find that for cetaceans sleep is characterized by USWS, a negligible amount or complete absence of rapid eye movement (REM) sleep, and a varying degree of movement during sleep associated with body size, and an asymmetrical eye state. We then compare the anatomy of the mammalian somnogenic system with what is known in cetaceans, highlighting areas where additional knowledge is needed to understand cetacean sleep. Three suggested functions of USWS (facilitation of movement, more efficient sensory processing and control of breathing) are discussed. Lastly, the possible selection pressures leading to this form of sleep are examined, leading us to the suggestion that the selection pressure necessitating the evolution of cetacean sleep was most likely the need to offset heat loss to the water from birth and throughout life. Aspects such as sentinel functions and breathing are likely to be proximate evolutionary phenomenon of this form of sleep.
Figures

References
-
- Amlaner CJ, Ball NJ, 1994. Avian sleep. In: Kryger MH, Roth T, Dement WC (Eds.), Principles and Practice of Sleep Medicine. second ed. Saunders, Philadelphia, pp. 81–94.
-
- Anderson PK, 1986. Dugongs of Shark Bay, Australia—seasonal migration, water temperature and forage. Natl. Geogr. Res 2, 473–490.
-
- Balance LT, 2002. Cetacean ecology. In: Perrin WF, Würsig B, Thewissen JGM (Eds.), Encyclopedia of Marine Mammals. Academic Press, New York, pp. 208–214.
-
- Bearzi G, Politi E, 1999. Diurnal behavior of free-ranging bottlenose dolphins in the Kvarneric (Northern Adriatic sea). Mar. Mamm. Sci 15, 1065–1097.
-
- Behrman G, 1990. The pineal organ (epiphysis cerebri) of the harbor porpoise (Phocoena phocoena, Linne, 1758). Aquat. Mamm 16, 96–100.
