

Structures of substrate- and inhibitor-bound adenosine deaminase from a human malaria parasite show a dramatic conformational change and shed light on drug selectivity
- PMID: 18602399
- PMCID: PMC2600493
- DOI: 10.1016/j.jmb.2008.06.048
Structures of substrate- and inhibitor-bound adenosine deaminase from a human malaria parasite show a dramatic conformational change and shed light on drug selectivity
Abstract
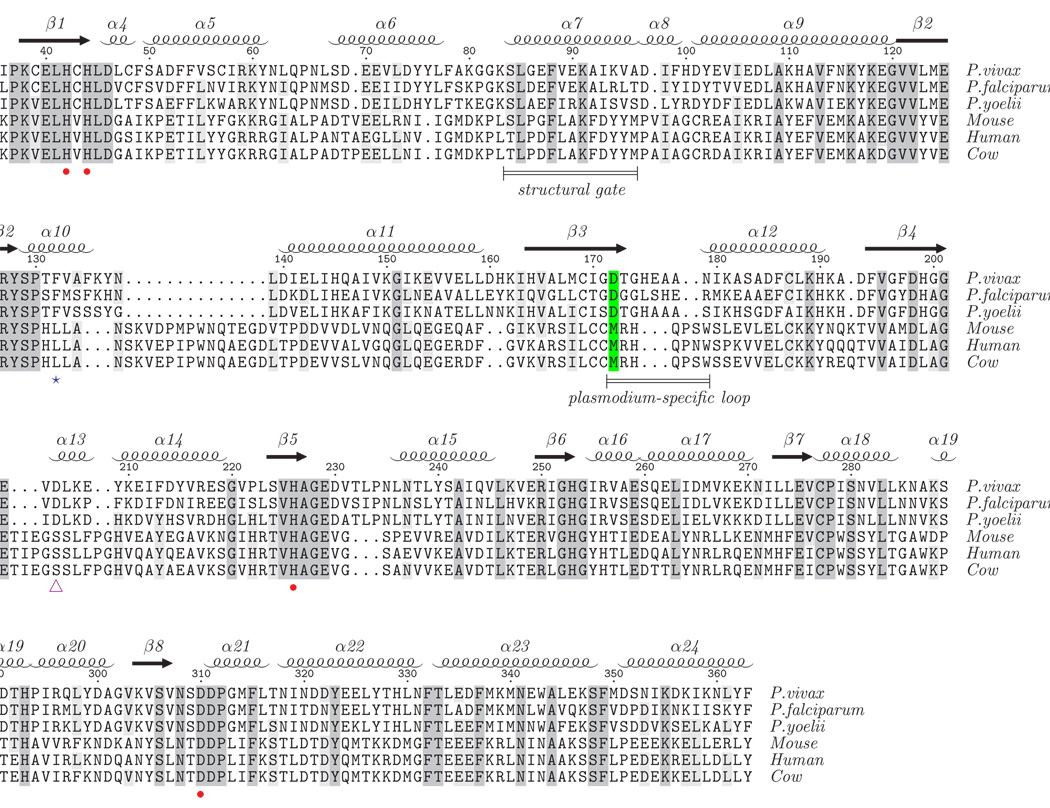
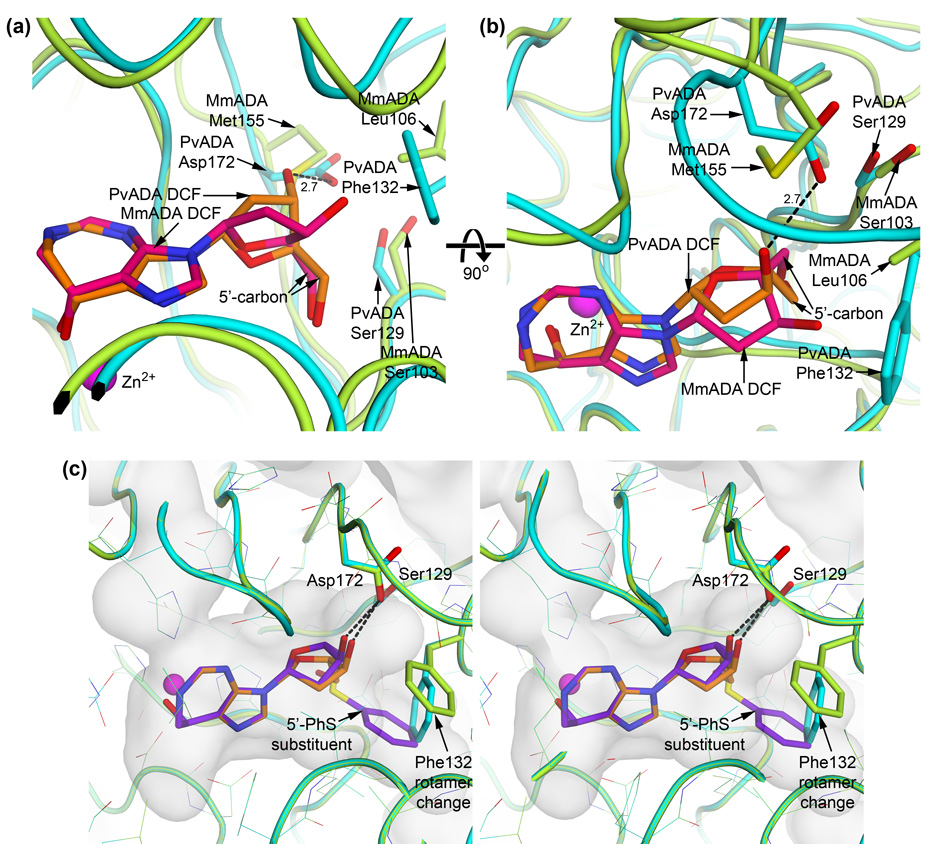
Plasmodium and other apicomplexan parasites are deficient in purine biosynthesis, relying instead on the salvage of purines from their host environment. Therefore, interference with the purine salvage pathway is an attractive therapeutic target. The plasmodial enzyme adenosine deaminase (ADA) plays a central role in purine salvage and, unlike mammalian ADA homologs, has a further secondary role in methylthiopurine recycling. For this reason, plasmodial ADA accepts a wider range of substrates, as it is responsible for deamination of both adenosine and 5'-methylthioadenosine. The latter substrate is not accepted by mammalian ADA homologs. The structural basis for this natural difference in specificity between plasmodial and mammalian ADA has not been well understood. We now report crystal structures of Plasmodium vivax ADA in complex with adenosine, guanosine, and the picomolar inhibitor 2'-deoxycoformycin. These structures highlight a drastic conformational change in plasmodial ADA upon substrate binding that has not been observed for mammalian ADA enzymes. Further, these complexes illuminate the structural basis for the differential substrate specificity and potential drug selectivity between mammalian and parasite enzymes.
Figures
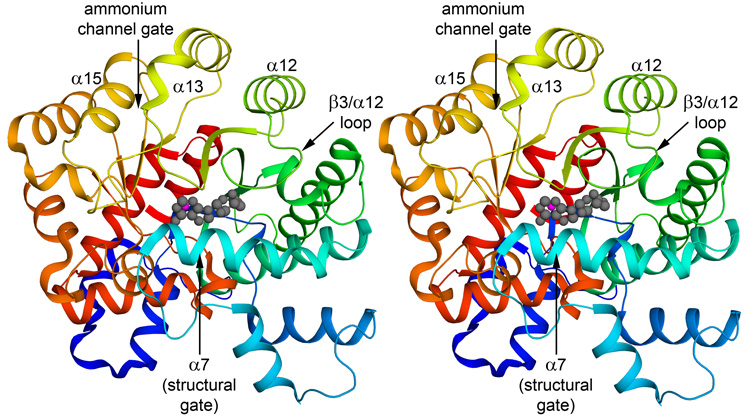
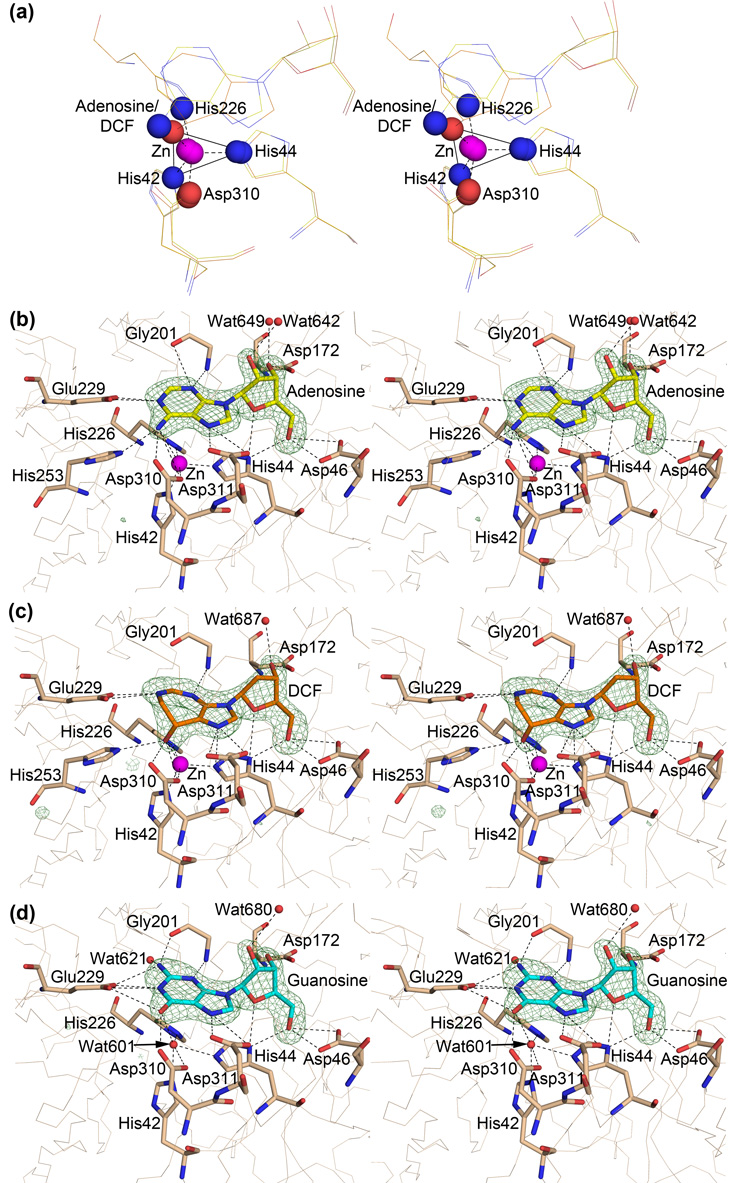
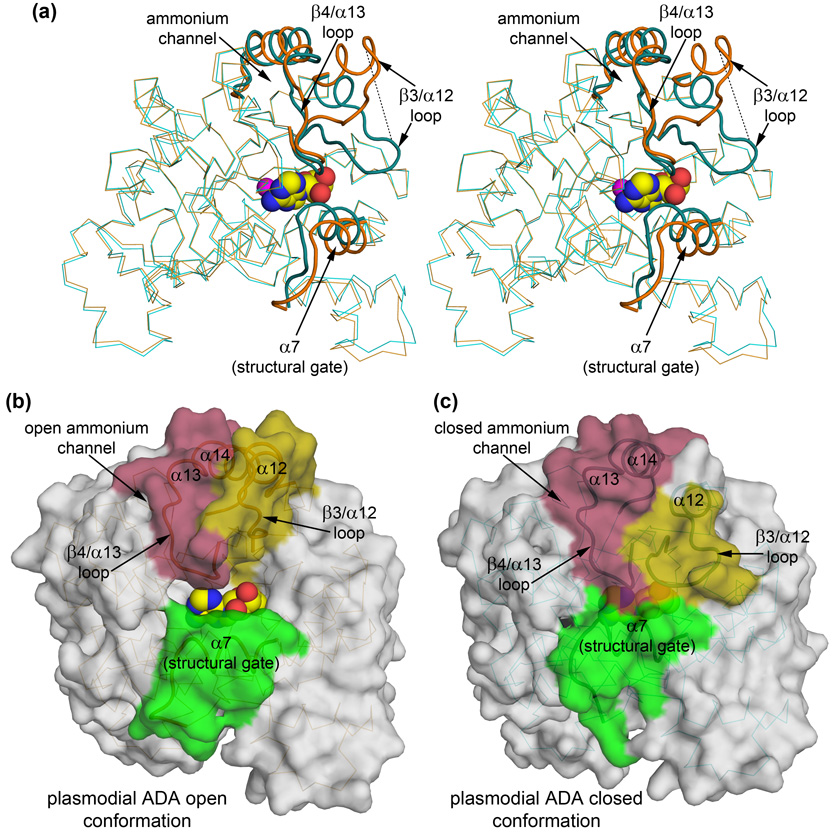
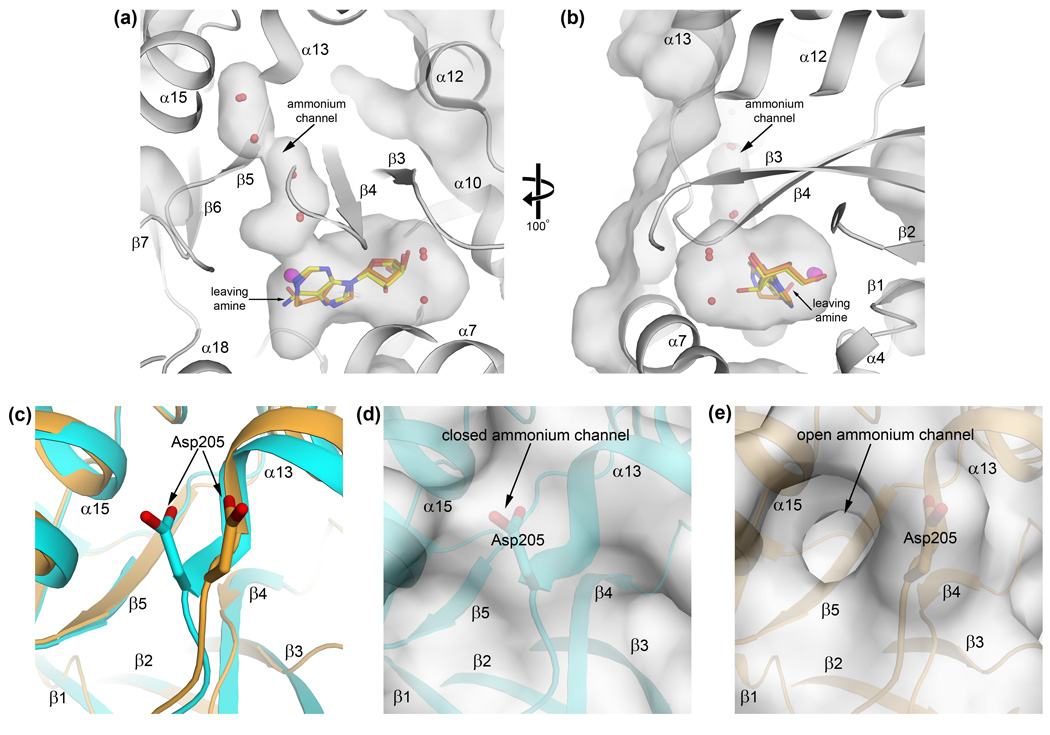
References
-
- Fan E, Baker D, Gelb MH, Buckner FS, Van Voorhis WC, Phizicky E, Dumont M, Mehlin C, Grayhack EJ, Sullivan M, Verlinde CL, DeTitta G, Meldrum D, Merritt EA, Earnest TN, Soltis M, Zucker F, Myler P, Schoenfeld L, Kim D, Worthey EA, LaCount D, Vignali M, Li J, Mondal S, Massey A, Carroll B, Gulde S, Luft JR, DeSoto L, Holl M, Caruthers JM, Bosch J, Robien MA, Arakaki T, Holmes MA, Le Trong I, Hol WG. Structural Genomics of Pathogenic Protozoa: An Overview. In: Bostjan K, Mitchell G, Huber T, editors. Methods in Molecular Biology. Vol. 426. 2008. pp. 497–513. - PubMed
-
- Kicska GA, Tyler PC, Evans GB, Furneaux RH, Kim K, Schramm VL. Transition State Analogue Inhibitors of Purine Nucleoside Phosphorylase from Plasmodium falciparum. J. Biol. Chem. 2002;277:3219–3225. - PubMed
-
- Kicska GA, Tyler PC, Evans GB, Furneaux RH, Schramm VL, Kim K. Purine-less Death in Plasmodium falciparum Induced by Immucillin-H, a Transition State Analogue of Purine Nucleoside Phosphorylase. J. Biol. Chem. 2002;277:3226–3231. - PubMed
-
- Ting L-M, Shi W, Lewandowicz A, Singh V, Mwakingwe A, Birck MR, Ringia EAT, Bench G, Madrid DC, Tyler PC, Evans GB, Furneaux RH, Schramm VL, Kim K. Targeting a Novel Plasmodium falciparum Purine Recycling Pathway with Specific Immucillins. J. Biol. Chem. 2005;280:9547–9554. - PubMed
-
- Webster HK, Wiesmann WP, Pavia CS. Adenosine deaminase in malaria infection: effect of 2'-deoxycoformycin in vivo. Advances in Experimental Medicine and Biology. 1984;165(Pt A):225–229. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
