

Novel modes of rhythmic burst firing at cognitively-relevant frequencies in thalamocortical neurons
- PMID: 18602904
- PMCID: PMC2778821
- DOI: 10.1016/j.brainres.2008.06.029
Novel modes of rhythmic burst firing at cognitively-relevant frequencies in thalamocortical neurons
Abstract
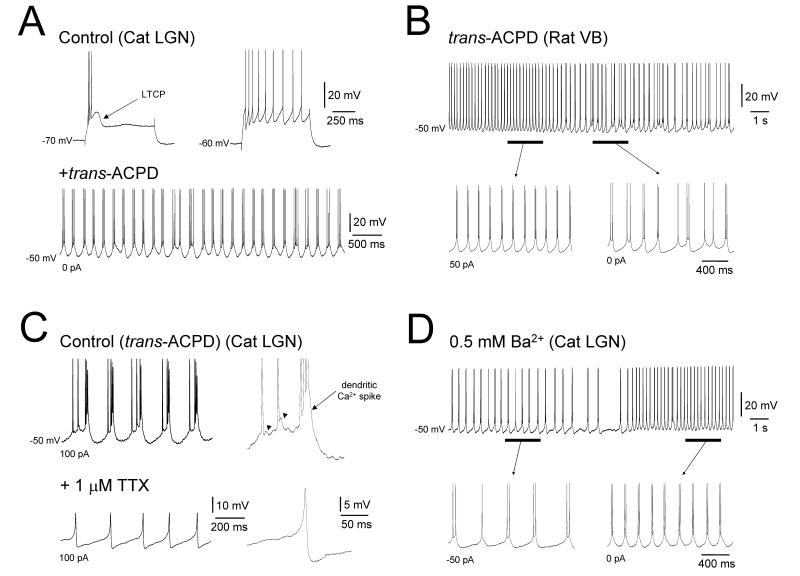
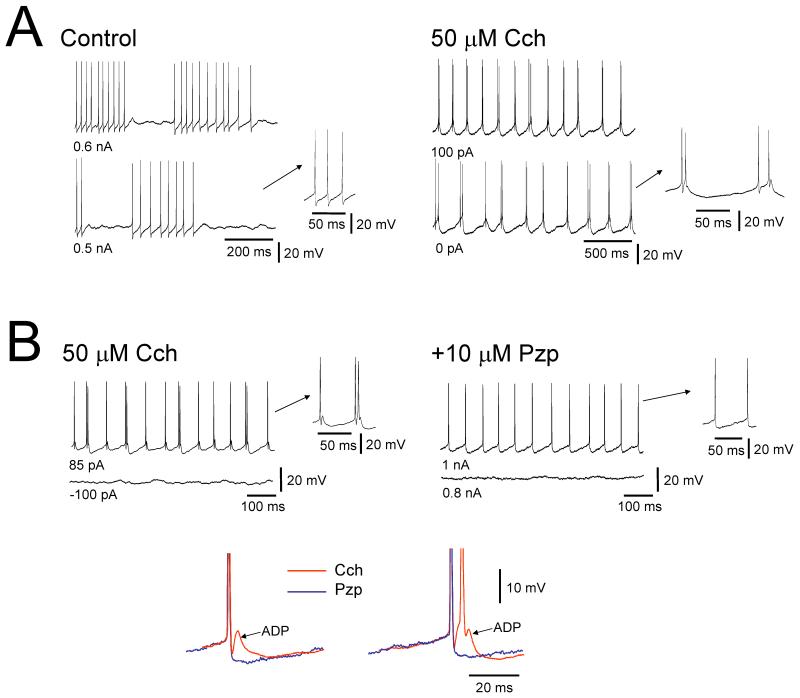
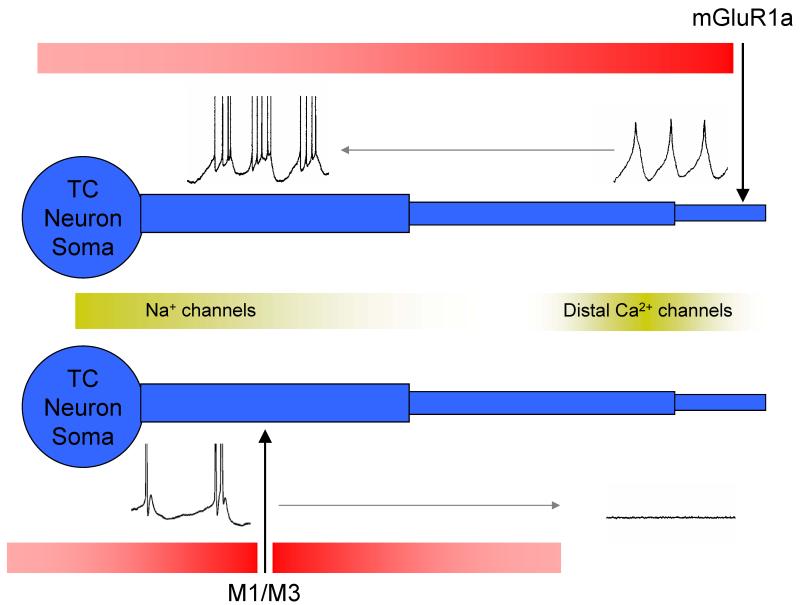
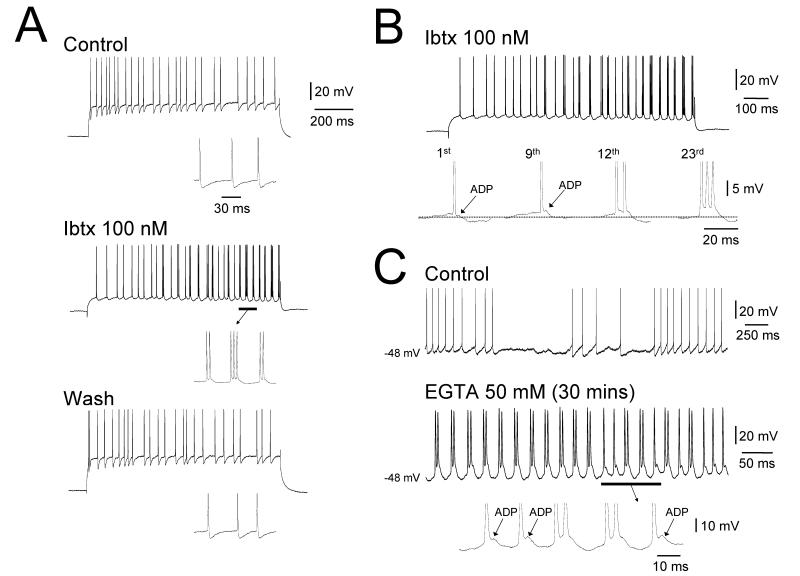
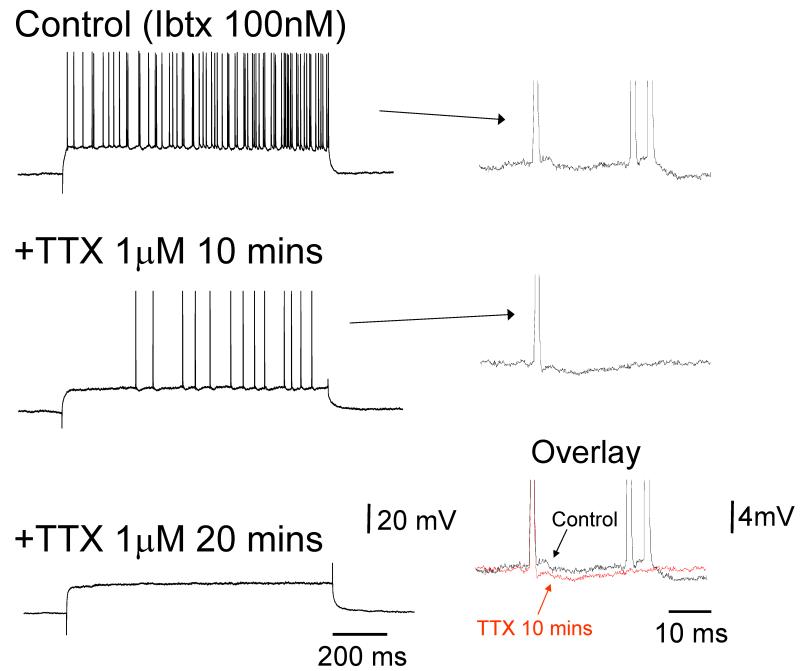
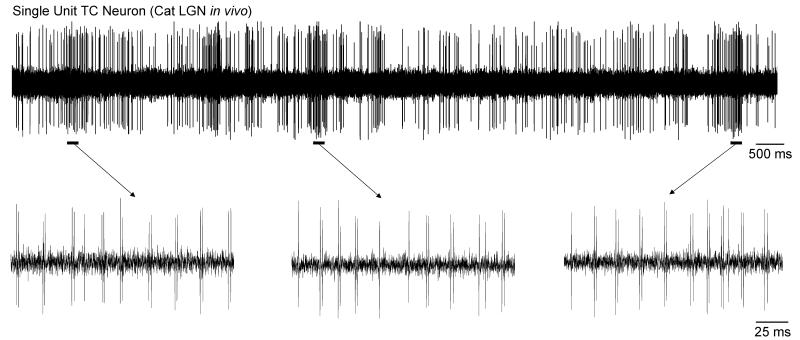
It is now widely accepted that certain types of cognitive functions are intimately related to synchronized neuronal oscillations at both low (alpha/theta) (4-7/8-13 Hz) and high (beta/gamma) (18-35/30-70 Hz) frequencies. The thalamus is a key participant in many of these oscillations, yet the cellular mechanisms by which this participation occurs are poorly understood. Here we describe how, under appropriate conditions, thalamocortical (TC) neurons from different nuclei can exhibit a wide array of largely unrecognised intrinsic oscillatory activities at a range of cognitively-relevant frequencies. For example, both metabotropic glutamate receptor (mGluR) and muscarinic Ach receptor (mAchR) activation can cause rhythmic bursting at alpha/theta frequencies. Interestingly, key differences exist between mGluR- and mAchR-induced bursting, with the former involving extensive dendritic Ca2+ electrogenesis and being mimicked by a non-specific block of K+ channels with Ba2+, whereas the latter appears to be more reliant on proximal Na+ channels and a prominent spike afterdepolarization (ADP). This likely relates to the differential somatodendritic distribution of mGluRs and mAChRs and may have important functional consequences. We also show here that in similarity to some neocortical neurons, inhibiting large-conductance Ca2+-activated K+ channels in TC neurons can lead to fast rhythmic bursting (FRB) at approximately 40 Hz. This activity also appears to rely on a Na+ channel-dependent spike ADP and may occur in vivo during natural wakefulness. Taken together, these results show that TC neurons are considerably more flexible than generally thought and strongly endorse a role for the thalamus in promoting a range of cognitively-relevant brain rhythms.
Figures
Similar articles
-
Medium-voltage 5-9-Hz oscillations give rise to spike-and-wave discharges in a genetic model of absence epilepsy: in vivo dual extracellular recording of thalamic relay and reticular neurons.Neuroscience. 2001;105(1):181-201. doi: 10.1016/s0306-4522(01)00182-8. Neuroscience. 2001. PMID: 11483311
-
Fast rhythmic bursting can be induced in layer 2/3 cortical neurons by enhancing persistent Na+ conductance or by blocking BK channels.J Neurophysiol. 2003 Feb;89(2):909-21. doi: 10.1152/jn.00573.2002. J Neurophysiol. 2003. PMID: 12574468
-
Simulation of GABAB-receptor-mediated K+ current in thalamocortical relay neurons: tonic firing, bursting, and oscillations.Biol Cybern. 1994;71(3):271-80. doi: 10.1007/BF00202766. Biol Cybern. 1994. PMID: 7918804
-
The role of T-channels in the generation of thalamocortical rhythms.CNS Neurol Disord Drug Targets. 2006 Dec;5(6):571-85. doi: 10.2174/187152706779025526. CNS Neurol Disord Drug Targets. 2006. PMID: 17168743 Review.
-
Bursting of thalamic neurons and states of vigilance.J Neurophysiol. 2006 Jun;95(6):3297-308. doi: 10.1152/jn.00166.2006. Epub 2006 Mar 22. J Neurophysiol. 2006. PMID: 16554502 Review.
Cited by
-
Alpha oscillations and the control of voluntary saccadic behavior.Exp Brain Res. 2012 Aug;221(2):123-8. doi: 10.1007/s00221-012-3167-8. Epub 2012 Jul 11. Exp Brain Res. 2012. PMID: 22782481 Free PMC article. Review.
-
Cortical Neural Synchronization Underlies Primary Visual Consciousness of Qualia: Evidence from Event-Related Potentials.Front Hum Neurosci. 2016 Jun 30;10:310. doi: 10.3389/fnhum.2016.00310. eCollection 2016. Front Hum Neurosci. 2016. PMID: 27445750 Free PMC article. Review.
-
Aberrant Network Activity in Schizophrenia.Trends Neurosci. 2017 Jun;40(6):371-382. doi: 10.1016/j.tins.2017.04.003. Epub 2017 May 14. Trends Neurosci. 2017. PMID: 28515010 Free PMC article. Review.
-
Infraslow (<0.1 Hz) oscillations in thalamic relay nuclei basic mechanisms and significance to health and disease states.Prog Brain Res. 2011;193:145-62. doi: 10.1016/B978-0-444-53839-0.00010-7. Prog Brain Res. 2011. PMID: 21854961 Free PMC article. Review.
-
Sensory and cortical activation of distinct glial cell subtypes in the somatosensory thalamus of young rats.Eur J Neurosci. 2010 Jul;32(1):29-40. doi: 10.1111/j.1460-9568.2010.07281.x. Eur J Neurosci. 2010. PMID: 20608967 Free PMC article.
References
-
- Adrian ED, Matthews BHC. The Berger Rhythm: potential changes from the occipital lobes in man. Brain. 1934;57:355–85. - PubMed
-
- Adrian ED, Yamagiwa K. The origin of the Berger rhythm. Brain. 1935;58:323–51.
-
- Anliker J. Variations in alpha voltage of the electroencephalogram and time perception. Science. 1963;140:1307–9. - PubMed
-
- Anliker J. Simultaneous changes in visual separation threshold and voltage of cortical alpha rhythm. Science. 1966;153:316–18. - PubMed
-
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat. Rev. Neurosci. 2007;8:45–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
