

Avoidance learning facilitates temporal processing in the primary auditory cortex
- PMID: 18603453
- PMCID: PMC2556562
- DOI: 10.1016/j.nlm.2008.05.003
Avoidance learning facilitates temporal processing in the primary auditory cortex
Abstract
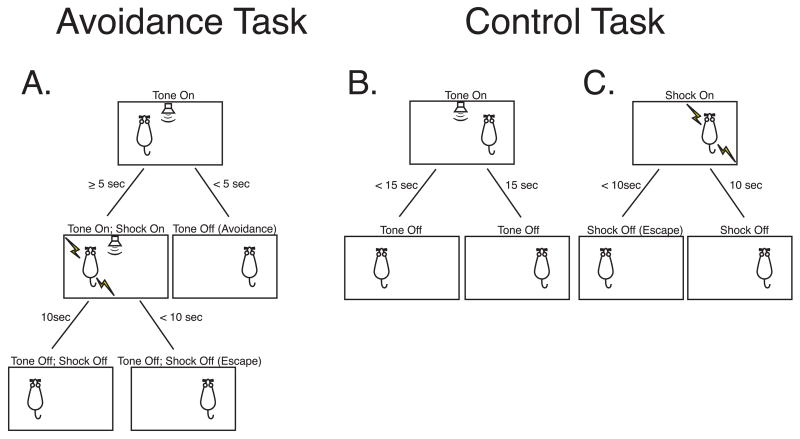
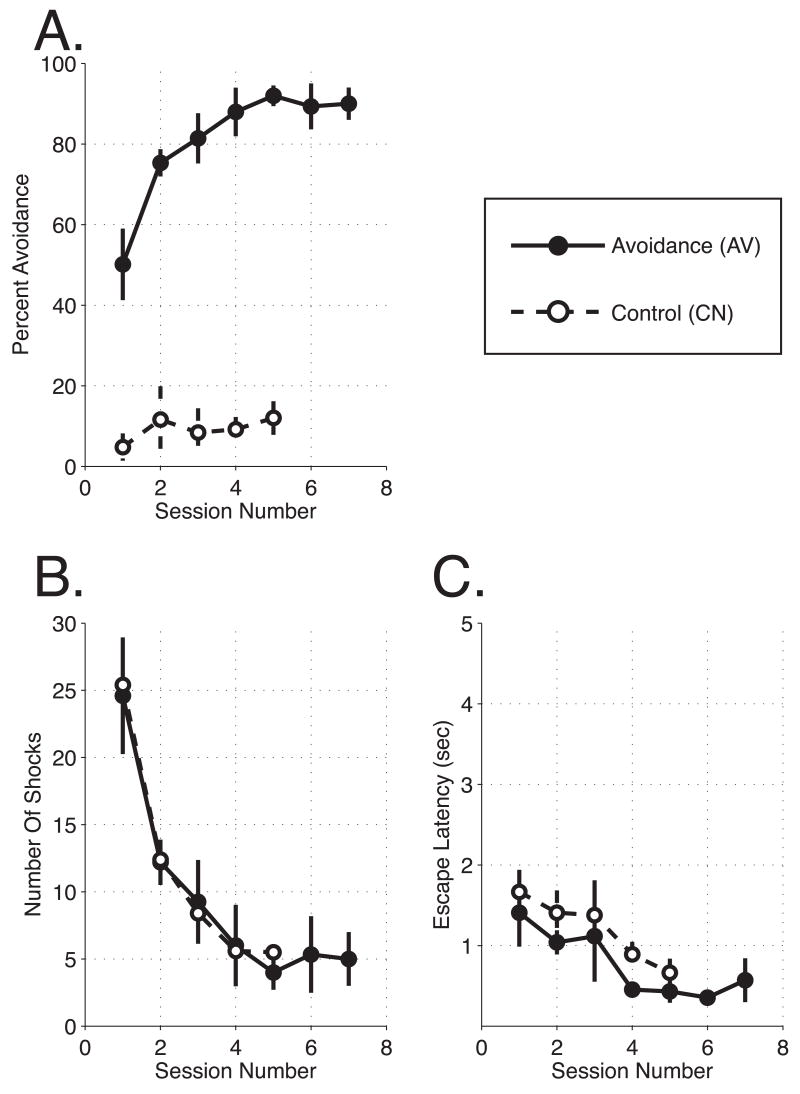
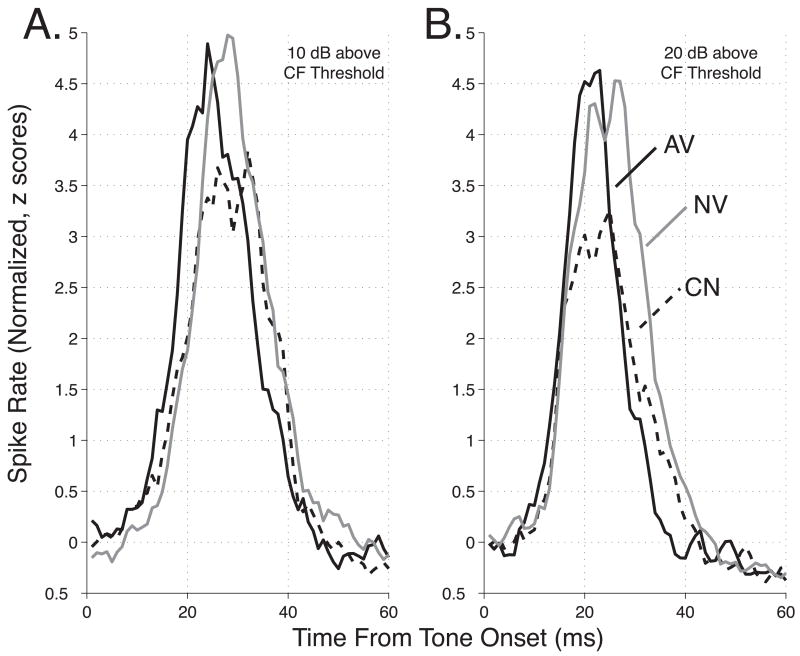
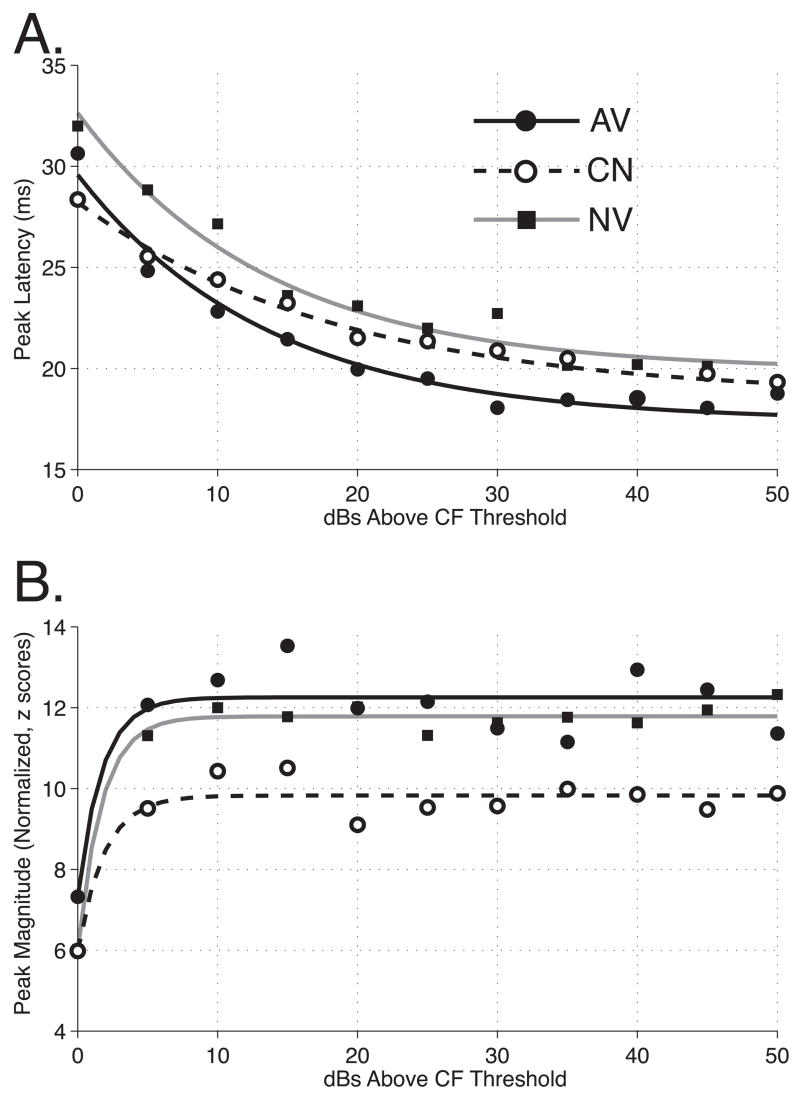
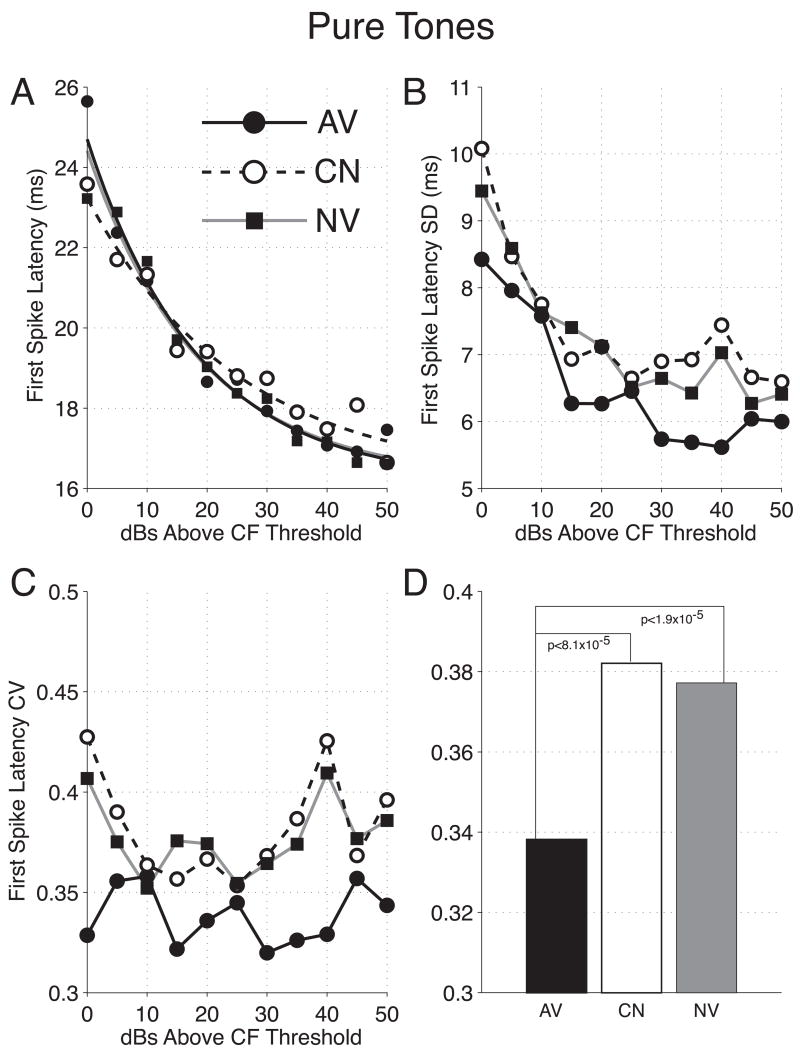
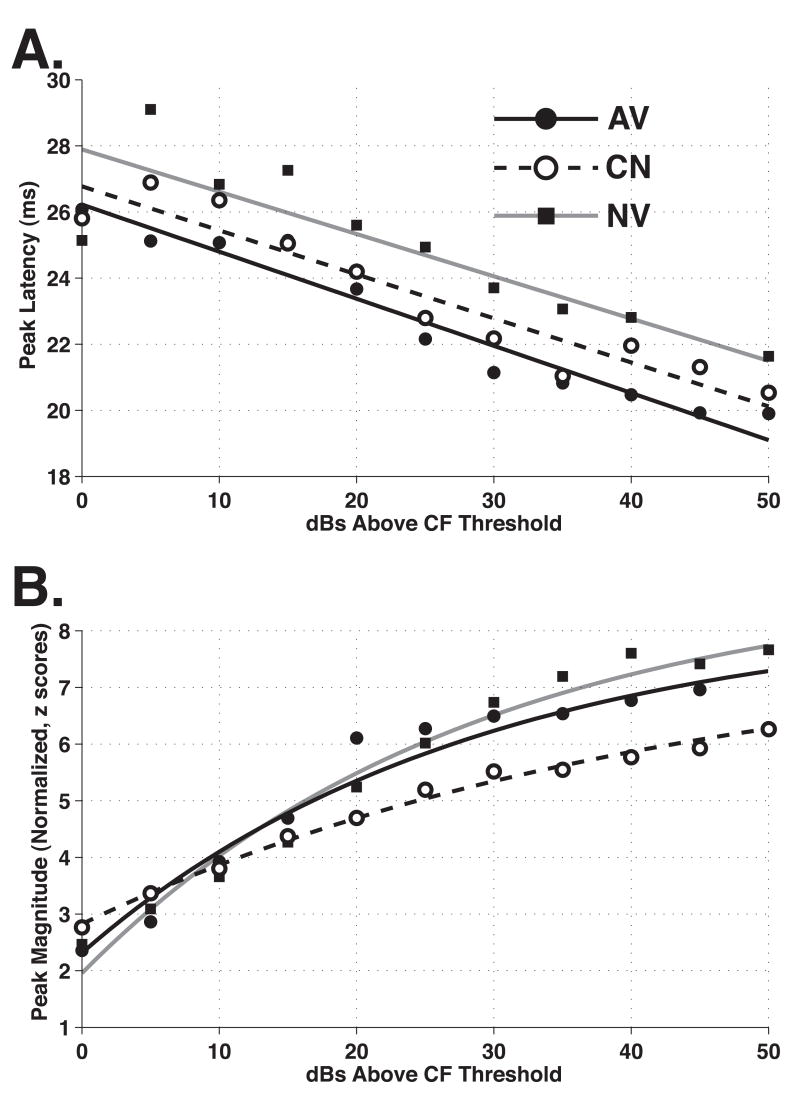
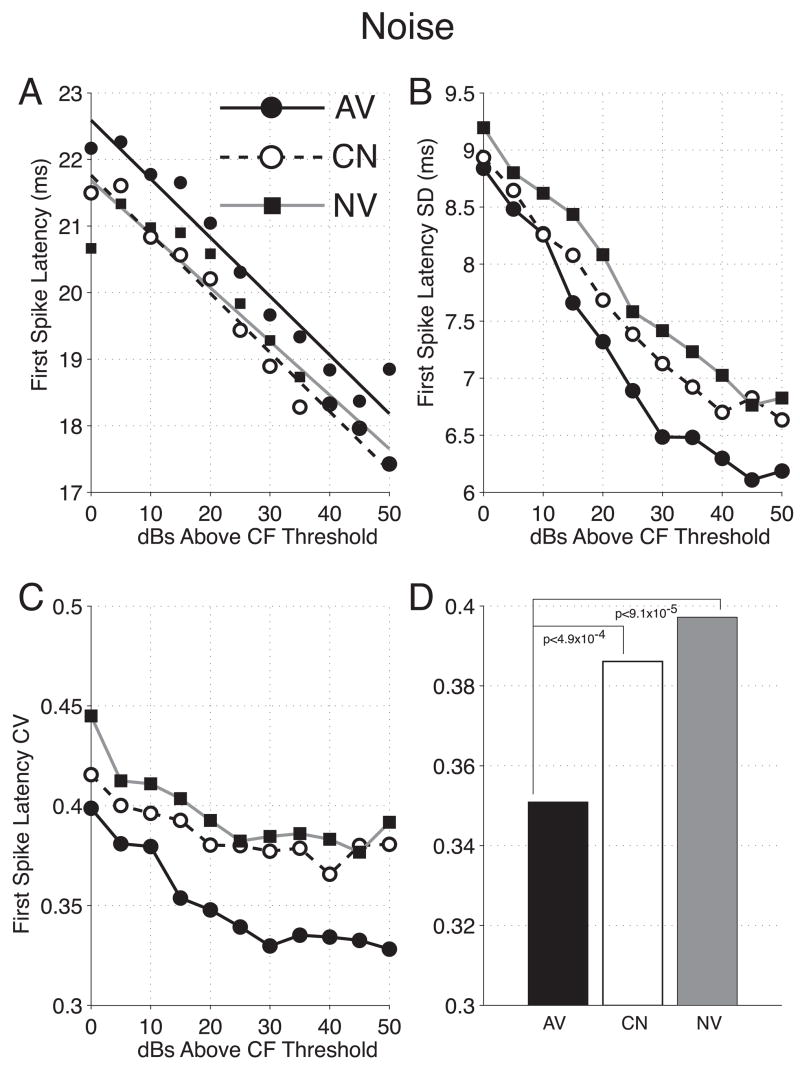
The primary auditory cortex is now known to be involved in learning and memory, as well as auditory perception. For example, spectral tuning often shifts toward or to the frequency of the conditioned stimulus during associative learning. As previous research has focused on tonal frequency, less is known about how learning might alter temporal parameters of response in the auditory cortex. This study addressed the effects of learning on the fidelity of temporal processing. Adult male rats were trained to avoid shock that was signaled by an 8.0 kHz tone. A novel control group received non-contingent tone and shock with shock probability decreasing over days to match the reduced number of shocks received by the avoidance group as they mastered the task. An untrained (nai ve) group served as a baseline. Following training, neuronal responses to white noise and a broad spectrum of tones were determined across the primary auditory cortex in a terminal experiment with subjects under general anesthesia. Avoidance conditioning significantly improved the precision of spike-timing: the coefficient of variation of 1st spike latency was significantly reduced in avoidance animals compared to controls and nai ves, both for tones and for noise. Additionally, avoidance learning was accompanied by a reduction of the latency of peak response, by 2.0-2.5 ms relative to nai ves and approximately 1.0 ms relative to controls. The shock-matched controls also exhibited significantly shorter peak latency of response than nai ves, demonstrating the importance of this non-avoidance control. Plasticity of temporal processing showed no evidence of frequency specificity and developed independently of the non-temporal parameters magnitude of response, frequency tuning and neural threshold, none of which were facilitated. The facilitation of temporal processing suggests that avoidance learning may increase synaptic strength either within the auditory cortex, in the subcortical auditory system, or both.
Figures
Similar articles
-
Induction of receptive field plasticity in the auditory cortex of the guinea pig during instrumental avoidance conditioning.Behav Neurosci. 1996 Oct;110(5):905-13. doi: 10.1037//0735-7044.110.5.905. Behav Neurosci. 1996. PMID: 8918994
-
Learning strategy trumps motivational level in determining learning-induced auditory cortical plasticity.Neurobiol Learn Mem. 2010 Feb;93(2):229-39. doi: 10.1016/j.nlm.2009.10.003. Epub 2009 Oct 21. Neurobiol Learn Mem. 2010. PMID: 19853056 Free PMC article.
-
CS-specific modifications of auditory evoked potentials in the behaviorally conditioned rat.Brain Res. 2017 Sep 1;1670:235-247. doi: 10.1016/j.brainres.2017.06.030. Epub 2017 Jun 30. Brain Res. 2017. PMID: 28673481
-
Physiological memory in primary auditory cortex: characteristics and mechanisms.Neurobiol Learn Mem. 1998 Jul-Sep;70(1-2):226-51. doi: 10.1006/nlme.1998.3850. Neurobiol Learn Mem. 1998. PMID: 9753599 Review.
-
Representation of acoustic events in the primary auditory cortex.J Exp Psychol Hum Percept Perform. 1993 Feb;19(1):203-16. doi: 10.1037//0096-1523.19.1.203. J Exp Psychol Hum Percept Perform. 1993. PMID: 8440986 Review.
Cited by
-
Memory Specific to Temporal Features of Sound Is Formed by Cue-Selective Enhancements in Temporal Coding Enabled by Inhibition of an Epigenetic Regulator.J Neurosci. 2021 Nov 3;41(44):9192-9209. doi: 10.1523/JNEUROSCI.0691-21.2021. Epub 2021 Sep 20. J Neurosci. 2021. PMID: 34544835 Free PMC article.
-
Spectral plasticity in monkey primary auditory cortex limits performance generalization in a temporal discrimination task.J Neurophysiol. 2020 Dec 1;124(6):1798-1814. doi: 10.1152/jn.00278.2020. Epub 2020 Sep 30. J Neurophysiol. 2020. PMID: 32997564 Free PMC article.
-
Viewpoints: Approaches to defining and investigating fear.Nat Neurosci. 2019 Aug;22(8):1205-1216. doi: 10.1038/s41593-019-0456-6. Nat Neurosci. 2019. PMID: 31332374 Free PMC article.
-
Behavioral training enhances cortical temporal processing in neonatally deafened juvenile cats.J Neurophysiol. 2011 Aug;106(2):944-59. doi: 10.1152/jn.00731.2010. Epub 2011 May 4. J Neurophysiol. 2011. PMID: 21543753 Free PMC article.
-
Representational gain in cortical area underlies increase of memory strength.Proc Natl Acad Sci U S A. 2010 Feb 23;107(8):3793-8. doi: 10.1073/pnas.1000159107. Epub 2010 Feb 4. Proc Natl Acad Sci U S A. 2010. PMID: 20133679 Free PMC article.
References
-
- Bakin JS, South DA, Weinberger NM. Induction of receptive field plasticity in the auditory cortex of the guinea pig during instrumental avoidance conditioning. Behavioral Neuroscience. 1996;110(5):905–913. - PubMed
-
- Bakin JS, Weinberger NM. Classical conditioning induces CS-specific receptive field plasticity in the auditory cortex of the guinea pig. Brain Research. 1990;536(1–2):271–286. - PubMed
-
- Bao S, Chan VT, Merzenich MM. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature. 2001;412(6842):79–83. - PubMed
-
- Bao S, Chang EF, Woods J, Merzenich MM. Temporal plasticity in the primary auditory cortex induced by operant perceptual learning. Nature Neuroscience. 2004;7(9):974–981. - PubMed
-
- Beitel RE, Schreiner CE, Cheung SW, Wang X, Merzenich MM. Reward-dependent plasticity in the primary auditory cortex of adult monkeys trained to discriminate temporally modulated signals. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(19):11070–11075. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
