

EphrinB1 controls cell-cell junctions through the Par polarity complex
- PMID: 18604196
- PMCID: PMC2614381
- DOI: 10.1038/ncb1758
EphrinB1 controls cell-cell junctions through the Par polarity complex
Abstract
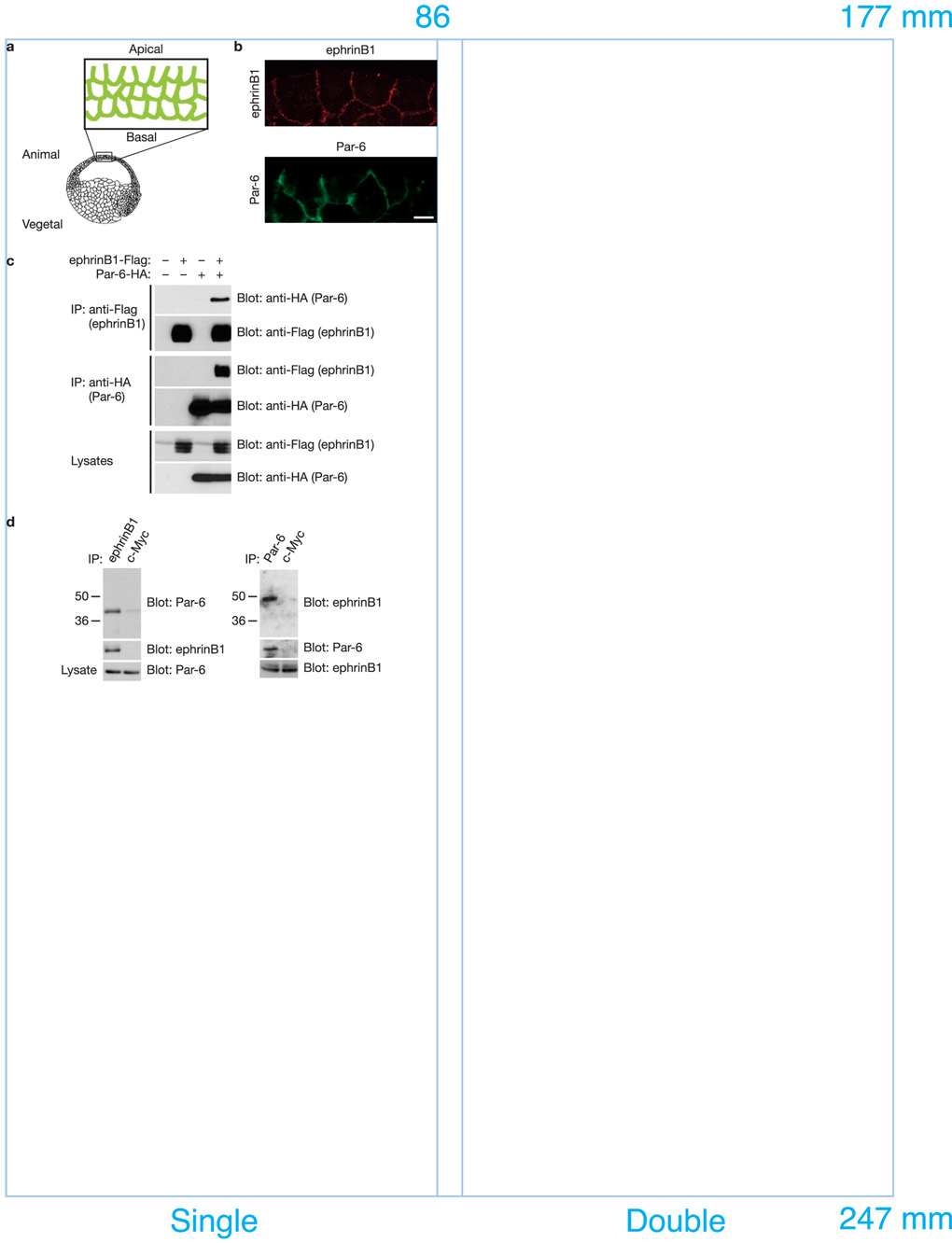
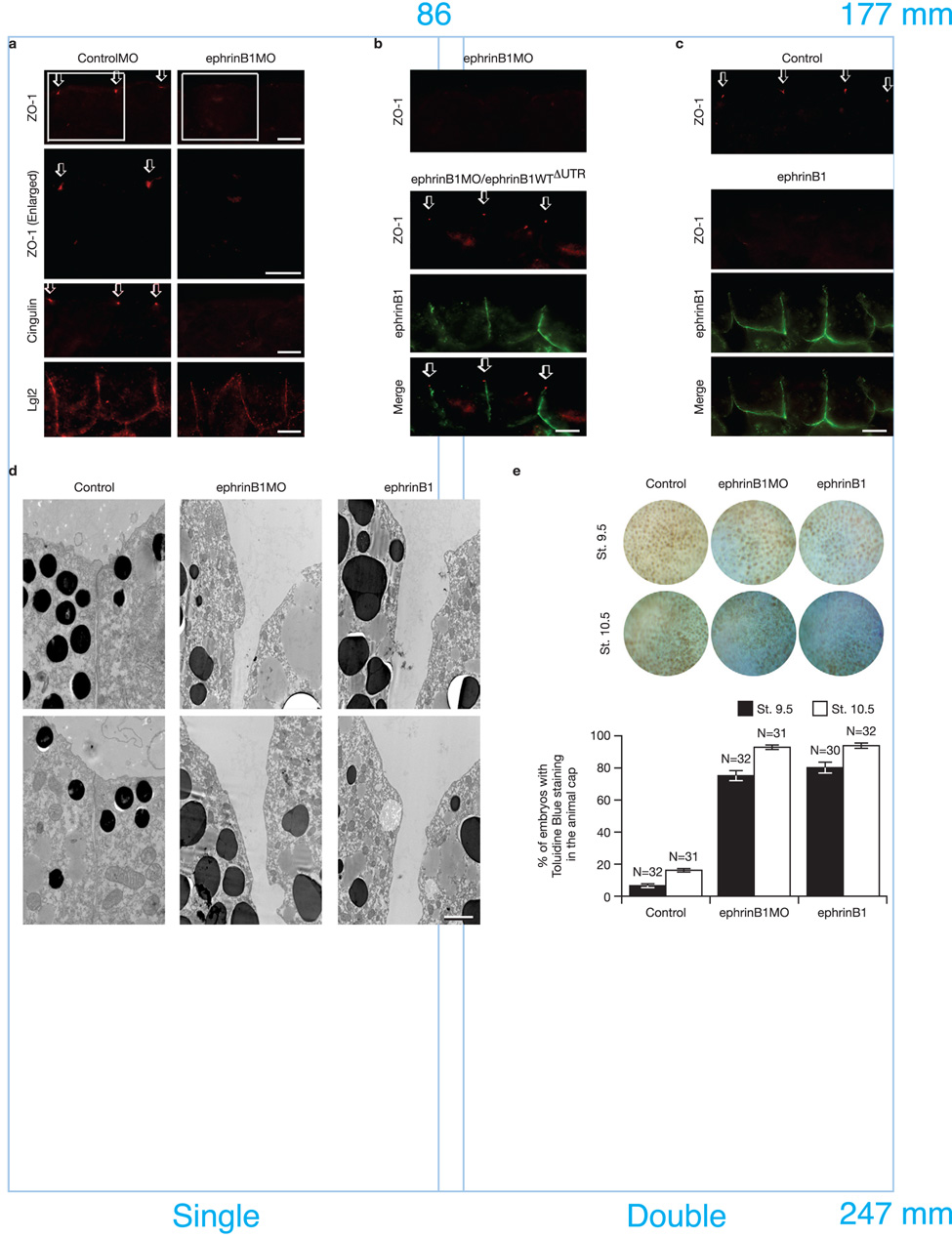
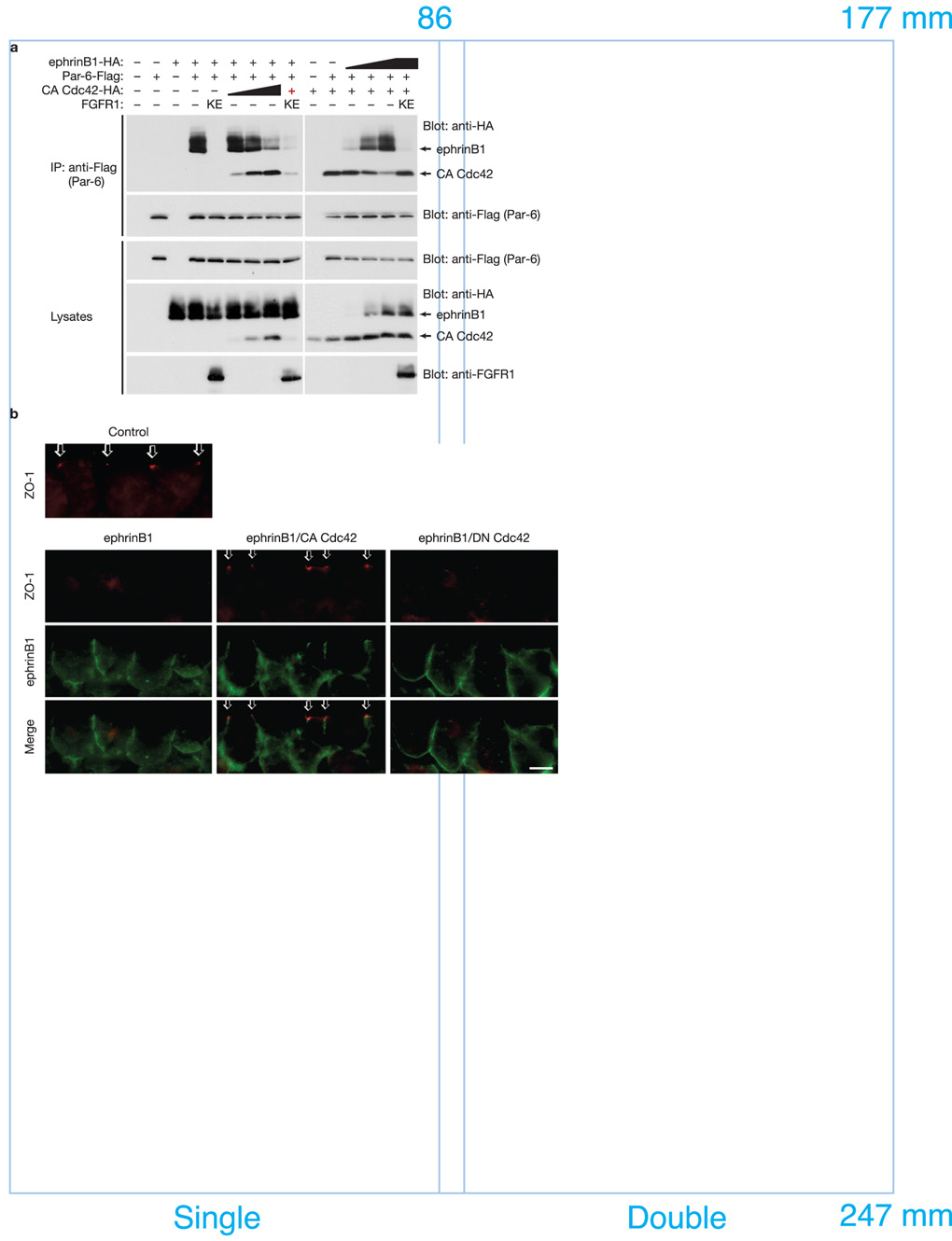
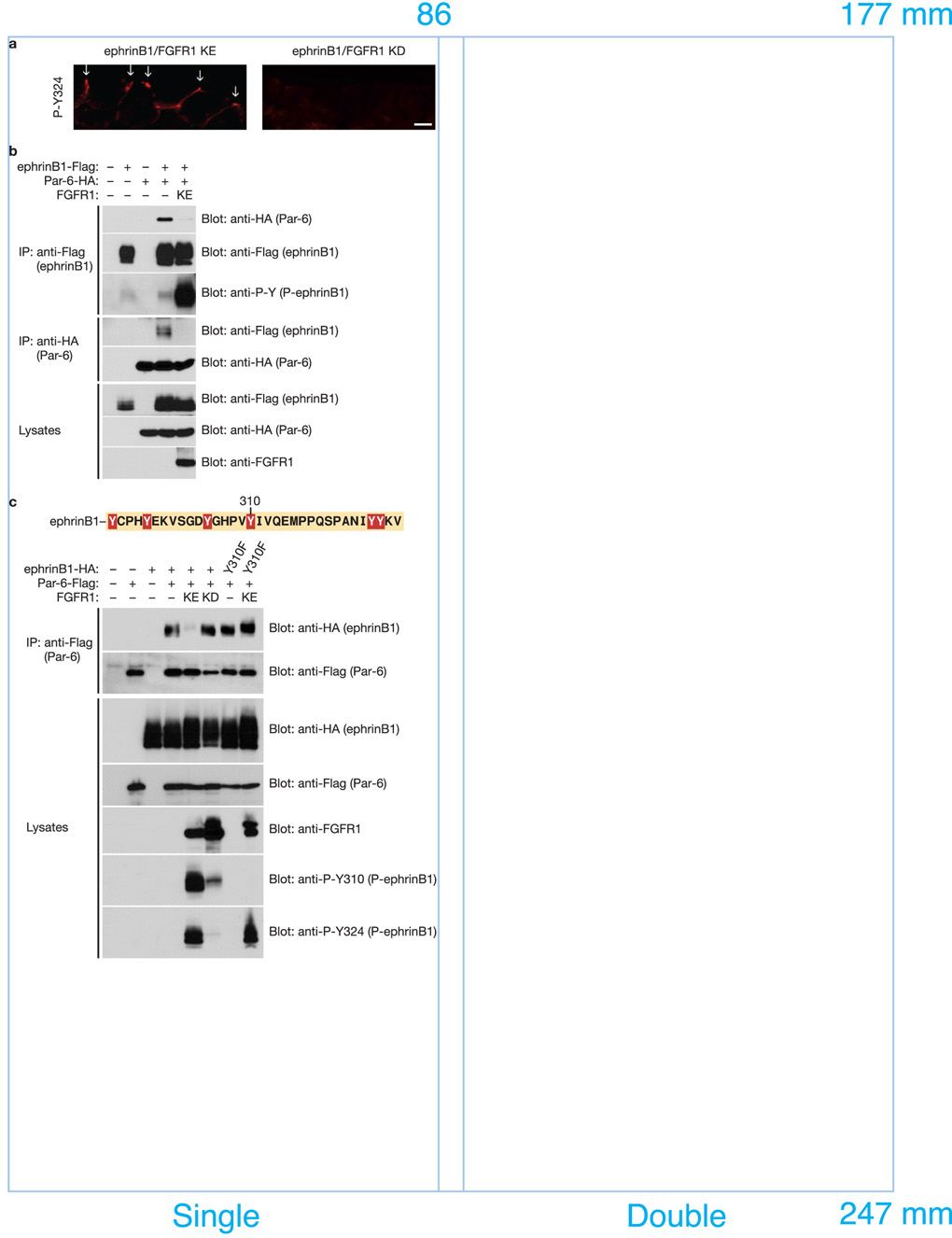
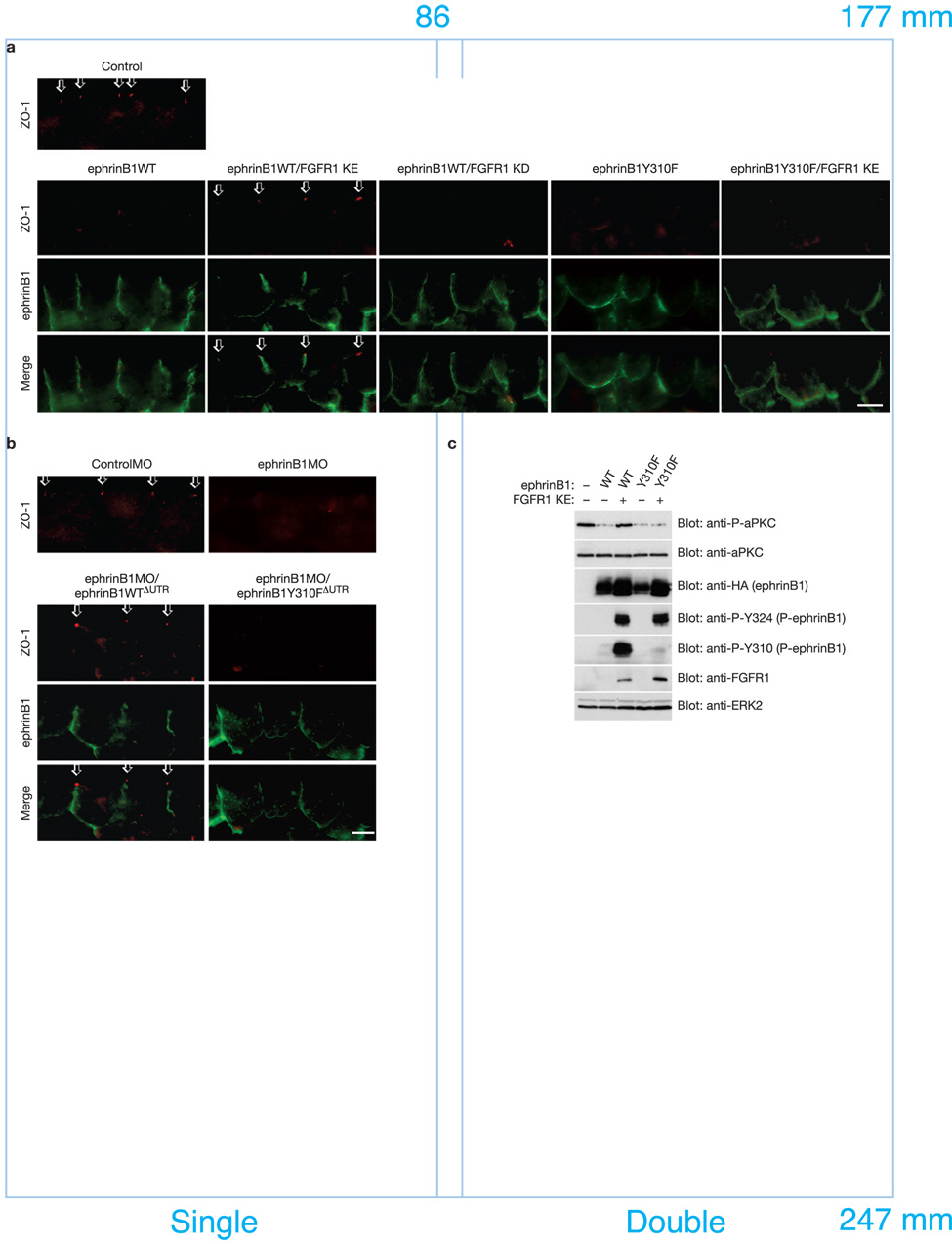
A body of evidence is emerging that shows a requirement for ephrin ligands in the proper migration of cells, and the formation of cell and tissue boundaries. These processes are dependent on the cell-cell adhesion system, which plays a crucial role in normal morphogenetic processes during development, as well as in invasion and metastasis. Although ephrinB ligands are bi-directional signalling molecules, the precise mechanism by which ephrinB1 signals through its intracellular domain to regulate cell-cell adhesion in epithelial cells remains unclear. Here, we present evidence that ephrinB1 associates with the Par polarity complex protein Par-6 (a scaffold protein required for establishing tight junctions) and can compete with the small GTPase Cdc42 for association with Par-6. This competition causes inactivation of the Par complex, resulting in the loss of tight junctions. Moreover, the interaction between ephrinB1 and Par-6 is disrupted by tyrosine phosphorylation of the intracellular domain of ephrinB1. Thus, we have identified a mechanism by which ephrinB1 signalling regulates cell-cell junctions in epithelial cells, and this may influence how we devise therapeutic interventions regarding these molecules in metastatic disease.
Figures
Similar articles
-
EphrinB reverse signaling in cell-cell adhesion: is it just par for the course?Cell Adh Migr. 2009 Jul-Sep;3(3):250-5. doi: 10.4161/cam.3.3.8211. Epub 2009 Jul 17. Cell Adh Migr. 2009. PMID: 19276658 Free PMC article.
-
Tyr-298 in ephrinB1 is critical for an interaction with the Grb4 adaptor protein.Biochem J. 2004 Jan 15;377(Pt 2):499-507. doi: 10.1042/BJ20031449. Biochem J. 2004. PMID: 14535844 Free PMC article.
-
Fibroblast growth factor receptor-induced phosphorylation of ephrinB1 modulates its interaction with Dishevelled.Mol Biol Cell. 2009 Jan;20(1):124-33. doi: 10.1091/mbc.e08-06-0662. Epub 2008 Nov 12. Mol Biol Cell. 2009. PMID: 19005214 Free PMC article.
-
The interdependence of the Rho GTPases and apicobasal cell polarity.Small GTPases. 2014;5(2):10. doi: 10.4161/21541248.2014.973768. Small GTPases. 2014. PMID: 25469537 Free PMC article. Review.
-
Epithelial junctions and Rho family GTPases: the zonular signalosome.Small GTPases. 2014;5(4):1-15. doi: 10.4161/21541248.2014.973760. Small GTPases. 2014. PMID: 25483301 Free PMC article. Review.
Cited by
-
Signaling pathways in cell polarity.Cold Spring Harb Perspect Biol. 2012 Jun 1;4(6):a009654. doi: 10.1101/cshperspect.a009654. Cold Spring Harb Perspect Biol. 2012. PMID: 22553378 Free PMC article.
-
Ligand targeting of EphA2 enhances keratinocyte adhesion and differentiation via desmoglein 1.Mol Biol Cell. 2010 Nov 15;21(22):3902-14. doi: 10.1091/mbc.E10-03-0242. Epub 2010 Sep 22. Mol Biol Cell. 2010. PMID: 20861311 Free PMC article.
-
Eph signaling in mitotic spindle orientation: what´s your angle here?Cell Cycle. 2019 Oct;18(20):2590-2597. doi: 10.1080/15384101.2019.1658479. Epub 2019 Sep 1. Cell Cycle. 2019. PMID: 31475621 Free PMC article.
-
Epithelial-mesenchymal transitions: the importance of changing cell state in development and disease.J Clin Invest. 2009 Jun;119(6):1438-49. doi: 10.1172/JCI38019. Epub 2009 Jun 1. J Clin Invest. 2009. PMID: 19487820 Free PMC article. Review.
-
The signaling adaptor GAB1 regulates cell polarity by acting as a PAR protein scaffold.Mol Cell. 2012 Aug 10;47(3):469-83. doi: 10.1016/j.molcel.2012.06.037. Mol Cell. 2012. PMID: 22883624 Free PMC article.
References
-
- Cortina C, et al. EphB-ephrin-B interactions suppress colorectal cancer progression by compartmentalizing tumor cells. Nat Genet. 2007;39:1376–1383. - PubMed
-
- Heroult M, Schaffner F, Augustin HG. Eph receptor and ephrin ligand-mediated interactions during angiogenesis and tumor progression. Exp Cell Res. 2006;312:642–650. - PubMed
-
- Zhang J, Hughes S. Role of the ephrin and Eph receptor tyrosine kinase families in angiogenesis and development of the cardiovascular system. J Pathol. 2006;208:453–461. - PubMed
-
- Wimmer-Kleikamp SH, Lackmann M. Eph-modulated cell morphology, adhesion and motility in carcinogenesis. IUBMB Life. 2005;57:421–431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
