

Light-sensitive neurons and channels mediate phototaxis in C. elegans
- PMID: 18604203
- PMCID: PMC2652401
- DOI: 10.1038/nn.2155
Light-sensitive neurons and channels mediate phototaxis in C. elegans
Abstract
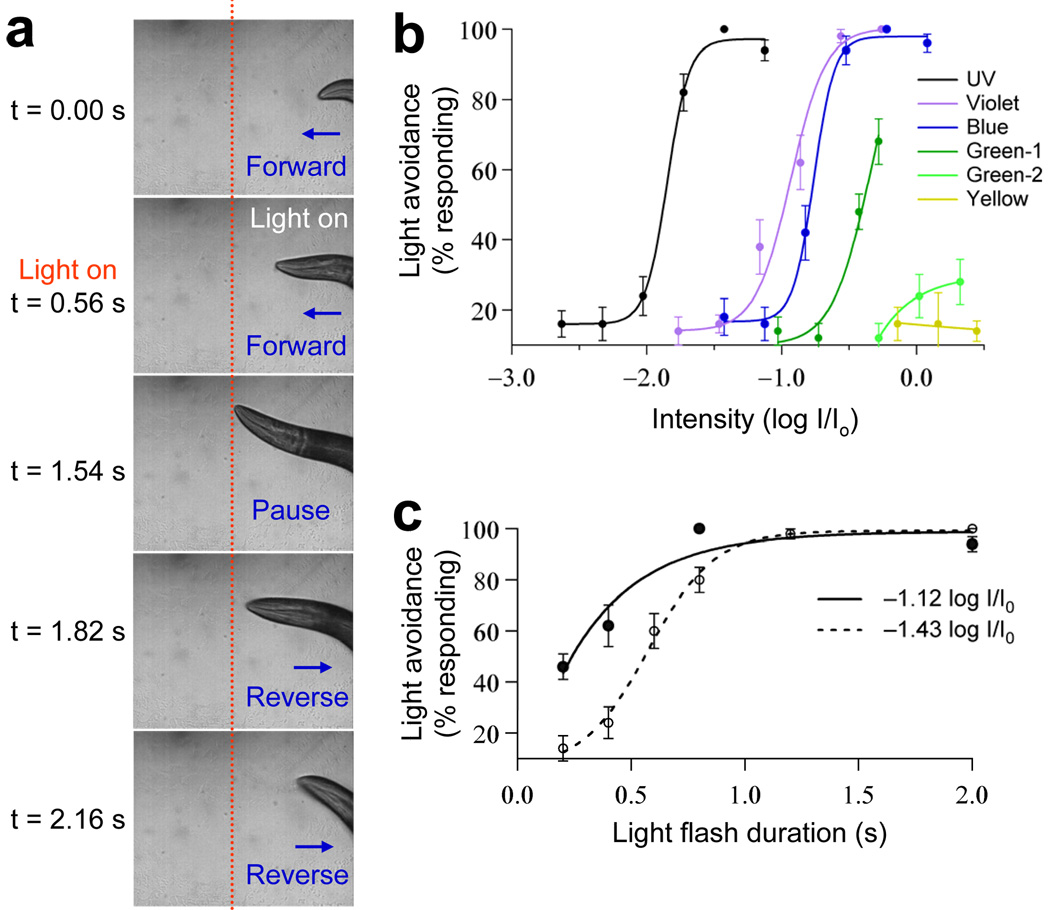
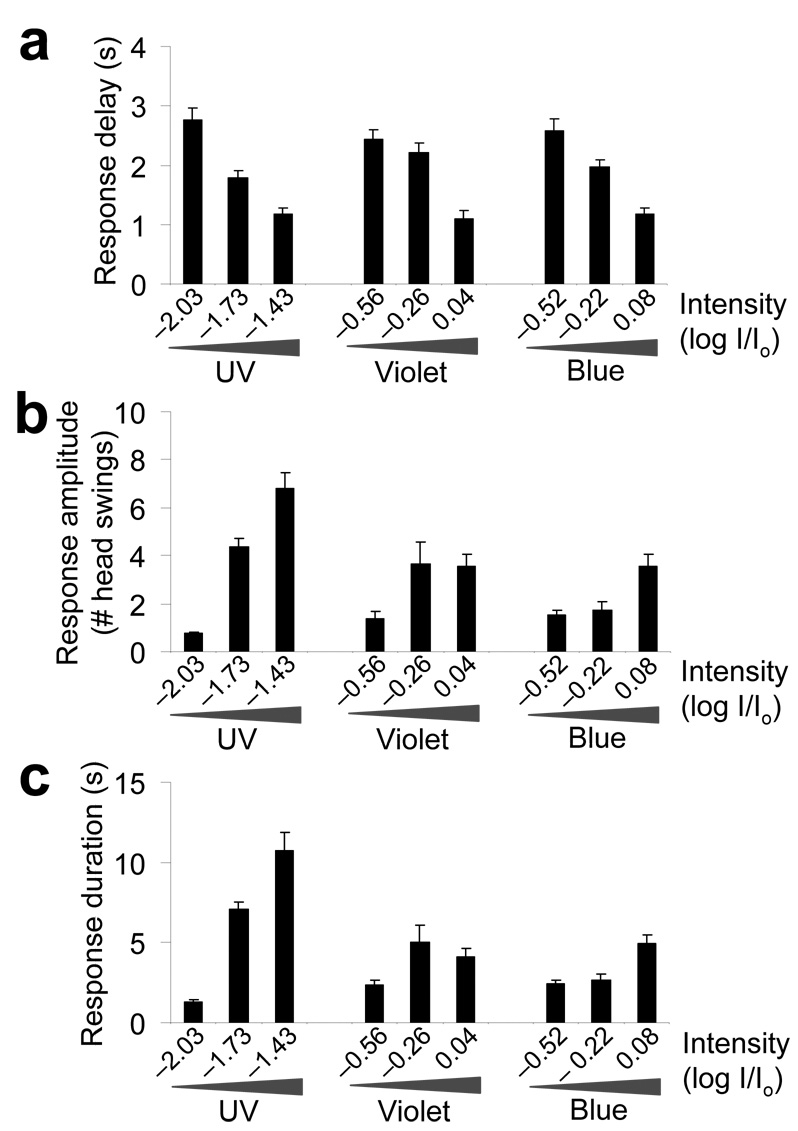
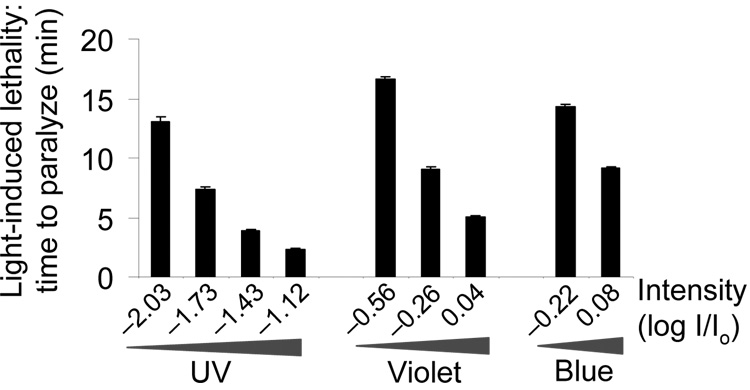
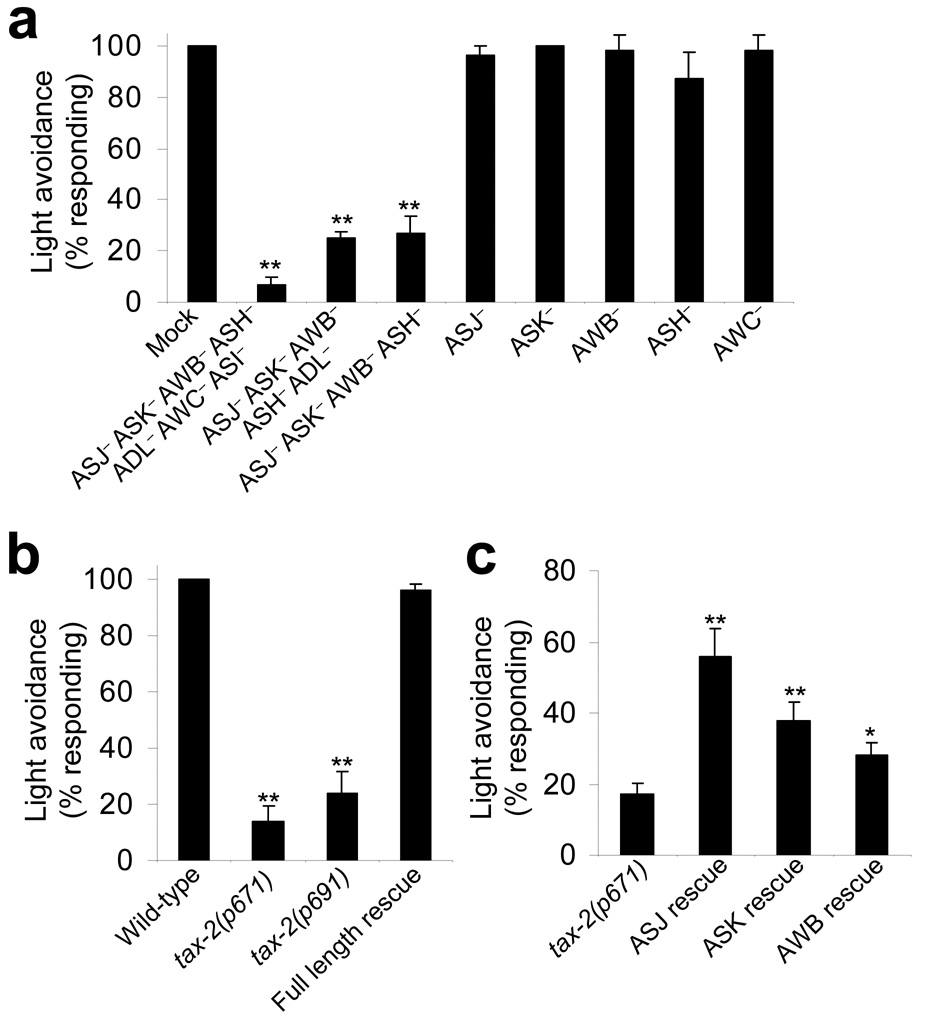
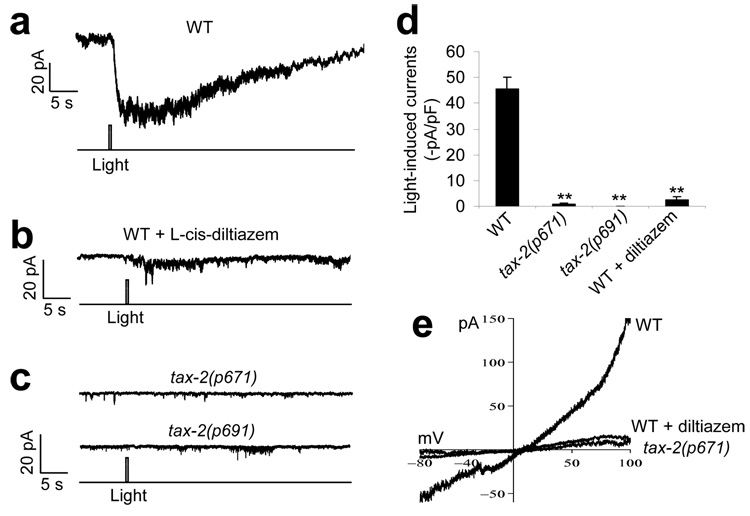
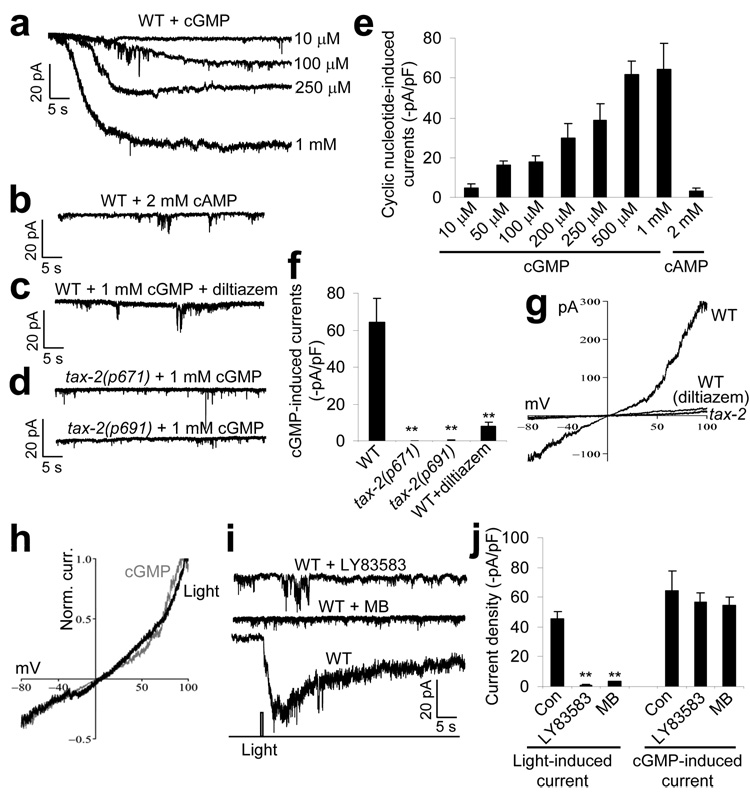
Phototaxis behavior is commonly observed in animals with light-sensing organs. C. elegans, however, is generally believed to lack phototaxis, as this animal lives in darkness (soil) and does not possess eyes. Here, we found that light stimuli elicited negative phototaxis in C. elegans and that this behavior is important for survival. We identified a group of ciliary sensory neurons as candidate photoreceptor cells for mediating phototaxis. Furthermore, we found that light excited photoreceptor cells by evoking a depolarizing conductance carried by cyclic guanosine monophosphate (cGMP)-sensitive cyclic nucleotide-gated (CNG) channels, revealing a conservation in phototransduction between worms and vertebrates. These results identify a new sensory modality in C. elegans and suggest that animals living in dark environments without light-sensing organs may not be presumed to be light insensitive. We propose that urbilaterians, the last common ancestor of bilaterians, might have already evolved a visual system that employs CNG channels and the second messenger cGMP for phototransduction.
Figures
Similar articles
-
C. elegans phototransduction requires a G protein-dependent cGMP pathway and a taste receptor homolog.Nat Neurosci. 2010 Jun;13(6):715-22. doi: 10.1038/nn.2540. Epub 2010 May 2. Nat Neurosci. 2010. PMID: 20436480 Free PMC article.
-
Defining specificity determinants of cGMP mediated gustatory sensory transduction in Caenorhabditis elegans.Genetics. 2013 Aug;194(4):885-901. doi: 10.1534/genetics.113.152660. Epub 2013 May 20. Genetics. 2013. PMID: 23695300 Free PMC article.
-
Optogenetic manipulation of cGMP in cells and animals by the tightly light-regulated guanylyl-cyclase opsin CyclOp.Nat Commun. 2015 Sep 8;6:8046. doi: 10.1038/ncomms9046. Nat Commun. 2015. PMID: 26345128 Free PMC article.
-
Modulation of cyclic-nucleotide-gated channels and regulation of vertebrate phototransduction.J Exp Biol. 2001 Sep;204(Pt 17):2921-31. doi: 10.1242/jeb.204.17.2921. J Exp Biol. 2001. PMID: 11551982 Review.
-
Tuning outer segment Ca2+ homeostasis to phototransduction in rods and cones.Adv Exp Med Biol. 2002;514:179-203. doi: 10.1007/978-1-4615-0121-3_11. Adv Exp Med Biol. 2002. PMID: 12596922 Review.
Cited by
-
Effect of pulse direct current signals on electrotactic movement of nematodes Caenorhabditis elegans and Caenorhabditis briggsae.Biomicrofluidics. 2011 Dec;5(4):44116-441169. doi: 10.1063/1.3665224. Epub 2011 Dec 15. Biomicrofluidics. 2011. PMID: 22232698 Free PMC article.
-
madd-4 plays a critical role in light against Bursaphelenchus xylophilus.Sci Rep. 2022 Aug 30;12(1):14796. doi: 10.1038/s41598-022-19263-9. Sci Rep. 2022. PMID: 36042283 Free PMC article.
-
Multimodal sensory processing in Caenorhabditis elegans.Open Biol. 2018 Jun;8(6):180049. doi: 10.1098/rsob.180049. Open Biol. 2018. PMID: 29925633 Free PMC article. Review.
-
Serotonin promotes exploitation in complex environments by accelerating decision-making.BMC Biol. 2016 Feb 4;14:9. doi: 10.1186/s12915-016-0232-y. BMC Biol. 2016. PMID: 26847342 Free PMC article.
-
Phototransduction motifs and variations.Cell. 2009 Oct 16;139(2):246-64. doi: 10.1016/j.cell.2009.09.029. Cell. 2009. PMID: 19837030 Free PMC article. Review.
References
-
- Kandel ER. The neurobiology of behavior. In: Kandel ER, Schwartz JH, Jessell TM, editors. Principles of Neural Science. McGraw-Hill Medical; 2000. pp. 5–66.
-
- Bargmann CI. Comparative chemosensation from receptors to ecology. Nature. 2006;444:295–301. CrossRef. - PubMed
-
- Wang T, Montell C. Phototransduction and retinal degeneration in Drosophila. Pflugers Arch. 2007;454:821–847. - PubMed
-
- Berson DM. Phototransduction in ganglion-cell photoreceptors. Pflugers Arch. 2007;454:849–855. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
