

Identification of neural outgrowth genes using genome-wide RNAi
- PMID: 18604272
- PMCID: PMC2435276
- DOI: 10.1371/journal.pgen.1000111
Identification of neural outgrowth genes using genome-wide RNAi
Abstract
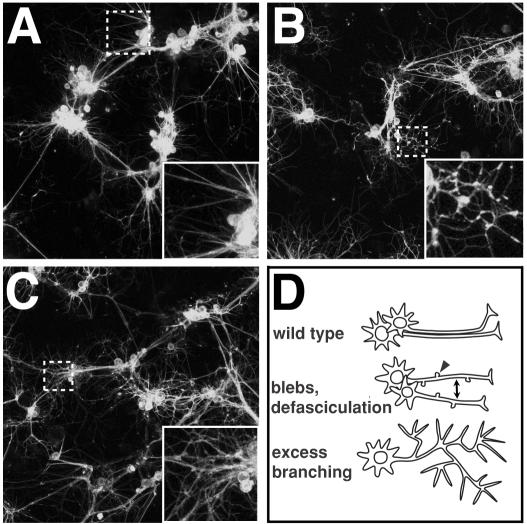
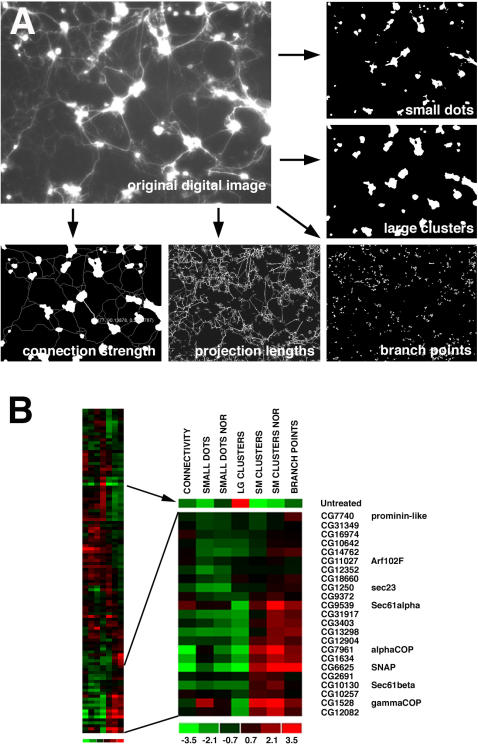
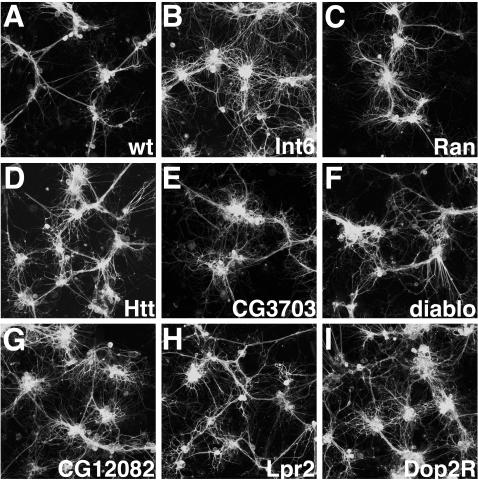
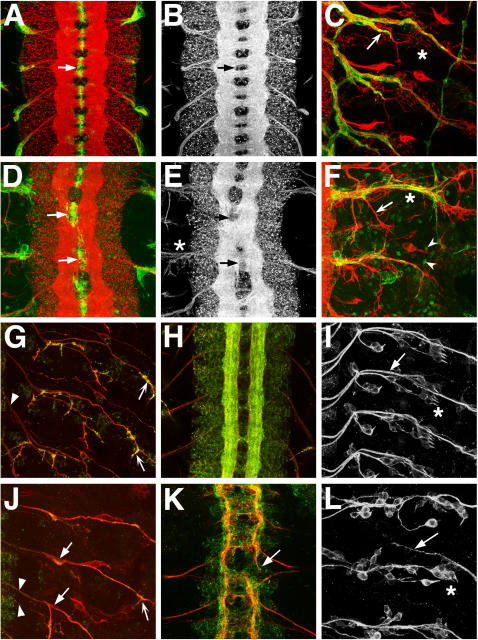
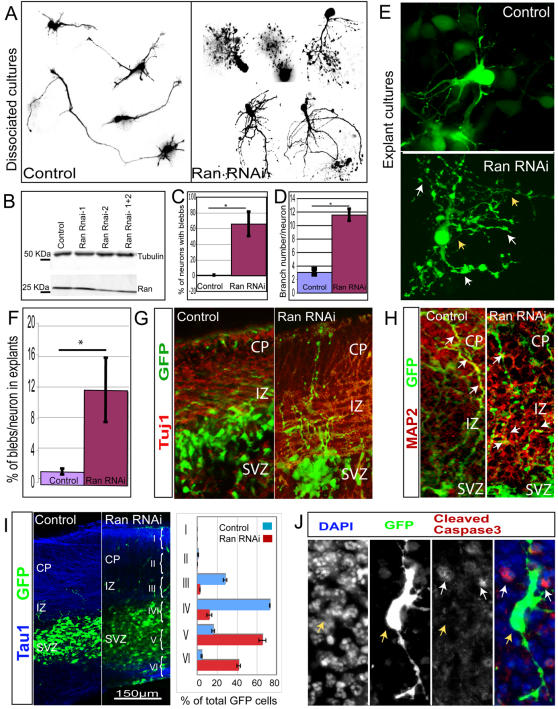
While genetic screens have identified many genes essential for neurite outgrowth, they have been limited in their ability to identify neural genes that also have earlier critical roles in the gastrula, or neural genes for which maternally contributed RNA compensates for gene mutations in the zygote. To address this, we developed methods to screen the Drosophila genome using RNA-interference (RNAi) on primary neural cells and present the results of the first full-genome RNAi screen in neurons. We used live-cell imaging and quantitative image analysis to characterize the morphological phenotypes of fluorescently labelled primary neurons and glia in response to RNAi-mediated gene knockdown. From the full genome screen, we focused our analysis on 104 evolutionarily conserved genes that when downregulated by RNAi, have morphological defects such as reduced axon extension, excessive branching, loss of fasciculation, and blebbing. To assist in the phenotypic analysis of the large data sets, we generated image analysis algorithms that could assess the statistical significance of the mutant phenotypes. The algorithms were essential for the analysis of the thousands of images generated by the screening process and will become a valuable tool for future genome-wide screens in primary neurons. Our analysis revealed unexpected, essential roles in neurite outgrowth for genes representing a wide range of functional categories including signalling molecules, enzymes, channels, receptors, and cytoskeletal proteins. We also found that genes known to be involved in protein and vesicle trafficking showed similar RNAi phenotypes. We confirmed phenotypes of the protein trafficking genes Sec61alpha and Ran GTPase using Drosophila embryo and mouse embryonic cerebral cortical neurons, respectively. Collectively, our results showed that RNAi phenotypes in primary neural culture can parallel in vivo phenotypes, and the screening technique can be used to identify many new genes that have important functions in the nervous system.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Schmid A, Schindelholz B, Zinn K. Combinatorial RNAi: a method for evaluating the functions of gene families in Drosophila. Trends Neurosci. 2002;25:71–74. - PubMed
-
- Mello CC, Conte D., Jr Revealing the world of RNA interference. Nature. 2004;431:338–342. - PubMed
-
- Tijsterman M, Plasterk RH. Dicers at RISC; the mechanism of RNAi. Cell. 2004;117:1–3. - PubMed
-
- Carpenter AE, Sabatini DM. Systematic genome-wide screens of gene function. Nat Rev Genet. 2004;5:11–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
