

Pervasive and persistent redundancy among duplicated genes in yeast
- PMID: 18604285
- PMCID: PMC2440806
- DOI: 10.1371/journal.pgen.1000113
Pervasive and persistent redundancy among duplicated genes in yeast
Abstract
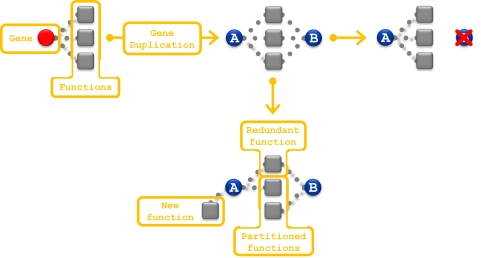
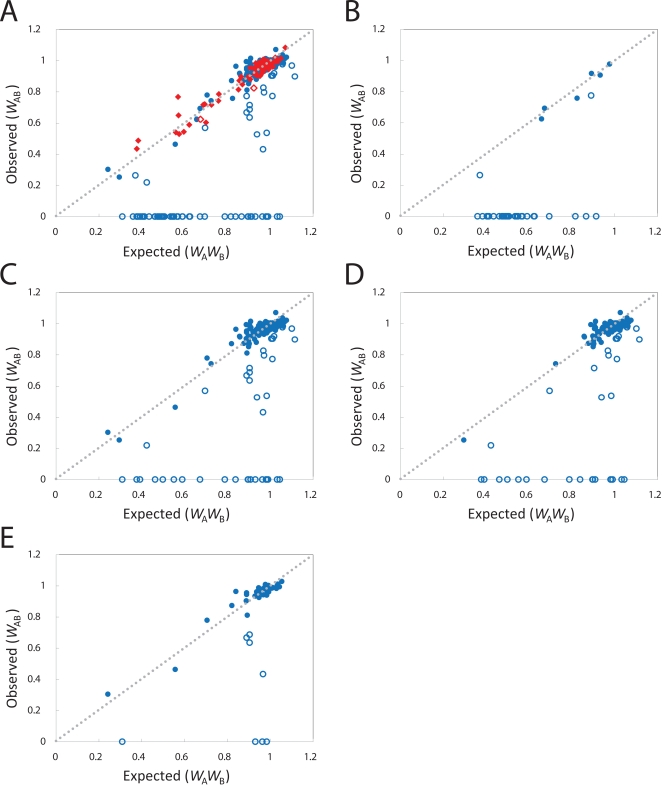
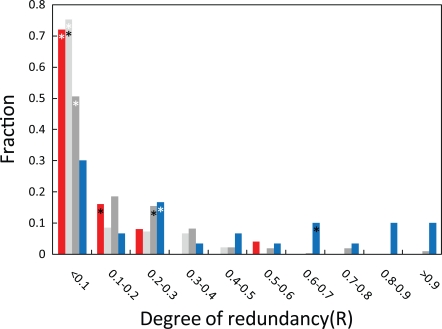
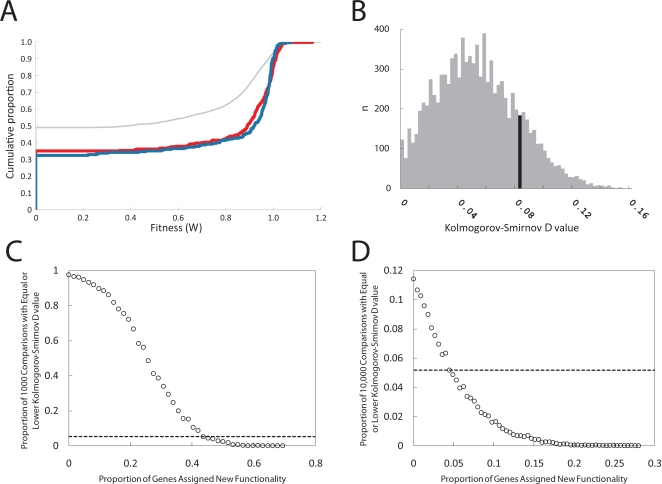
The loss of functional redundancy is the key process in the evolution of duplicated genes. Here we systematically assess the extent of functional redundancy among a large set of duplicated genes in Saccharomyces cerevisiae. We quantify growth rate in rich medium for a large number of S. cerevisiae strains that carry single and double deletions of duplicated and singleton genes. We demonstrate that duplicated genes can maintain substantial redundancy for extensive periods of time following duplication ( approximately 100 million years). We find high levels of redundancy among genes duplicated both via the whole genome duplication and via smaller scale duplications. Further, we see no evidence that two duplicated genes together contribute to fitness in rich medium substantially beyond that of their ancestral progenitor gene. We argue that duplicate genes do not often evolve to behave like singleton genes even after very long periods of time.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ohno S. Evolution by gene duplication. Berlin, New York: Springer-Verlag; 1970. p. xv, 160.
-
- Gu Z, Steinmetz LM, Gu X, Scharfe C, Davis RW, et al. Role of duplicate genes in genetic robustness against null mutations. Nature. 2003;421:63–66. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
