

SUMOylation of the MAGUK protein CASK regulates dendritic spinogenesis
- PMID: 18606847
- PMCID: PMC2447900
- DOI: 10.1083/jcb.200712094
SUMOylation of the MAGUK protein CASK regulates dendritic spinogenesis
Abstract
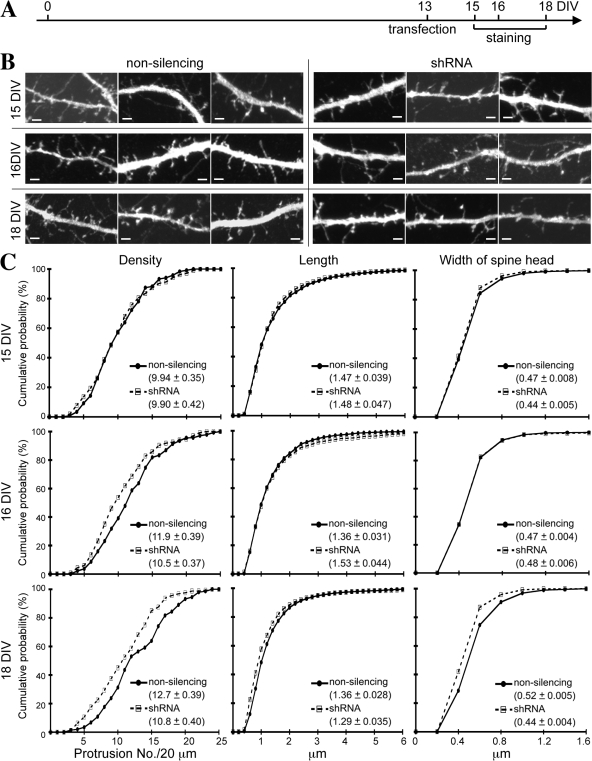
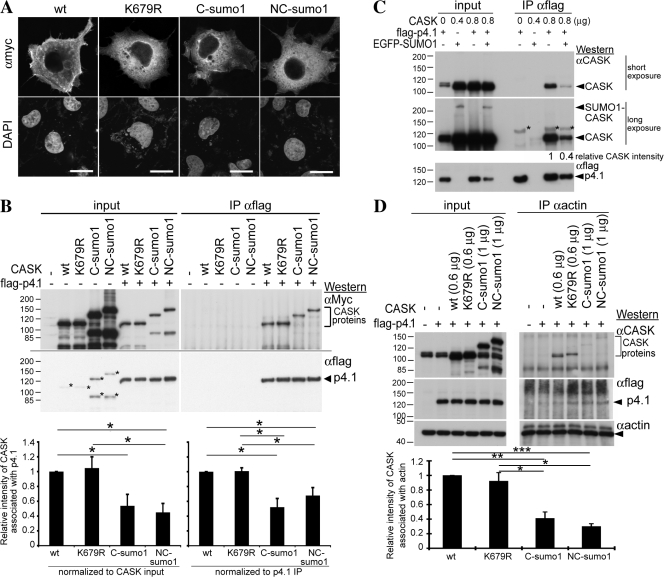
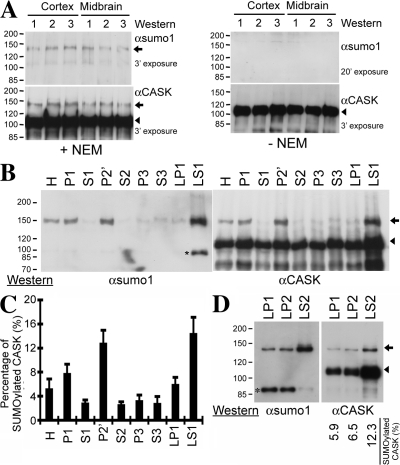
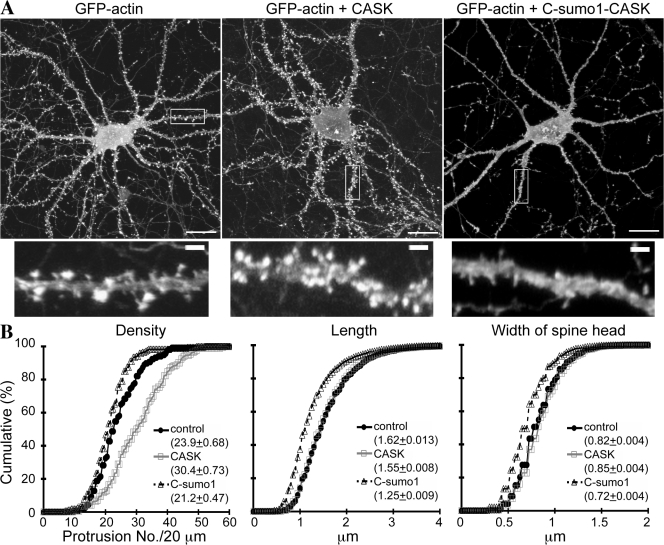
Membrane-associated guanylate kinase (MAGUK) proteins interact with several synaptogenesis-triggering adhesion molecules. However, direct evidence for the involvement of MAGUK proteins in synapse formation is lacking. In this study, we investigate the function of calcium/calmodulin-dependent serine protein kinase (CASK), a MAGUK protein, in dendritic spine formation by RNA interference. Knockdown of CASK in cultured hippocampal neurons reduces spine density and shrinks dendritic spines. Our analysis of the time course of RNA interference and CASK overexpression experiments further suggests that CASK stabilizes or maintains spine morphology. Experiments using only the CASK PDZ domain or a mutant lacking the protein 4.1-binding site indicate an involvement of CASK in linking transmembrane adhesion molecules and the actin cytoskeleton. We also find that CASK is SUMOylated. Conjugation of small ubiquitin-like modifier 1 (SUMO1) to CASK reduces the interaction between CASK and protein 4.1. Overexpression of a CASK-SUMO1 fusion construct, which mimicks CASK SUMOylation, impairs spine formation. Our study suggests that CASK contributes to spinogenesis and that this is controlled by SUMOylation.
Figures




References
-
- Alkuraya, F.S., I. Saadi, J.J. Lund, A. Turbe-Doan, C.C. Morton, and R.L. Maas. 2006. SUMO1 haploinsufficiency leads to cleft lip and palate. Science. 313:1751. - PubMed
-
- Biederer, T., and T.C. Sudhof. 2001. CASK and protein 4.1 support F-actin nucleation on neurexins. J. Biol. Chem. 276:47869–47876. - PubMed
-
- Biederer, T., Y. Sara, M. Mozhayeva, D. Atasoy, X. Liu, E.T. Kavalali, and T.C. Sudhof. 2002. SynCAM, a synaptic adhesion molecule that drives synapse assembly. Science. 297:1525–1531. - PubMed
-
- Borg, J.P., S.W. Straight, S.M. Kaech, M. de Taddeo-Borg, D.E. Kroon, D. Karnak, R.S. Turner, S.K. Kim, and B. Margolis. 1998. Identification of an evolutionarily conserved heterotrimeric protein complex involved in protein targeting. J. Biol. Chem. 273:31633–31636. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
