

Review
doi: 10.1016/j.str.2008.05.007.
Protein-protein interactions in the membrane: sequence, structural, and biological motifs
Affiliations
- PMID: 18611372
- PMCID: PMC3771515
- DOI: 10.1016/j.str.2008.05.007
Item in Clipboard
Review
Protein-protein interactions in the membrane: sequence, structural, and biological motifs
Structure.
2008 Jul.
Abstract
Single-span transmembrane (TM) helices have structural and functional roles well beyond serving as mere anchors to tether water-soluble domains in the vicinity of the membrane. They frequently direct the assembly of protein complexes and mediate signal transduction in ways analogous to small modular domains in water-soluble proteins. This review highlights different sequence and structural motifs that direct TM assembly and discusses their roles in diverse biological processes. We believe that TM interactions are potential therapeutic targets, as evidenced by natural proteins that modulate other TM interactions and recent developments in the design of TM-targeting peptides.
Figures
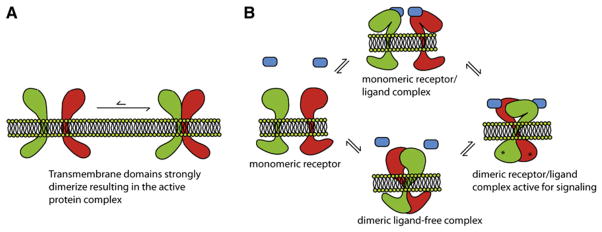
(A) Interacting TM helices can form static associations in the membrane or (B) participate in regulated switchable interactions in a dynamic equilibrium that regulates signaling. A simplified schematic of the coupled equilibria involving a dimeric receptor-ligand complex is depicted; however, signal transduction can potentially travel in both directions across the membrane. The receptor is in equilibrium between the monomeric and dimeric states, and ligand can bind to either. Along the bottom path, ligand binding to a dimeric receptor leads to a conformational change and signal transduction across the membrane. On the top path, the binding of ligand to a monomeric receptor can induce dimerization of the receptor and the subsequent conformational changes. Depending on the local ligand concentration and the expression level of the receptor in the membrane, different cell types and tissues can potentially fine-tune their responses as needed.
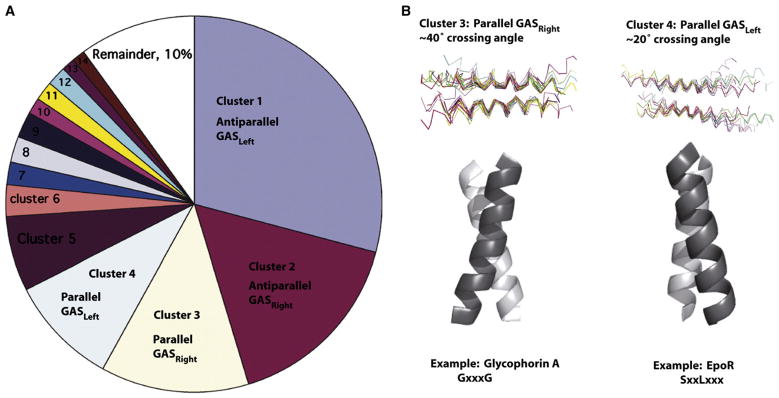
(A) Geometric clustering of interacting TM pairs in the crystallographic database by rmsd places two-thirds of TM helices into four categories: parallel or antiparallel GASLeft, and parallel or antiparallel GASRight (Walters and DeGrado, 2006). (B) The majority of known interacting monotypic TMs fall into parallel clusters. Parallel GASRight pairs have a ~40° crossing angle and a GX3G-like motif. Parallel GASLeft helices have a ~20° crossing angle and often a heptad repeat of small residues (G/A/S).
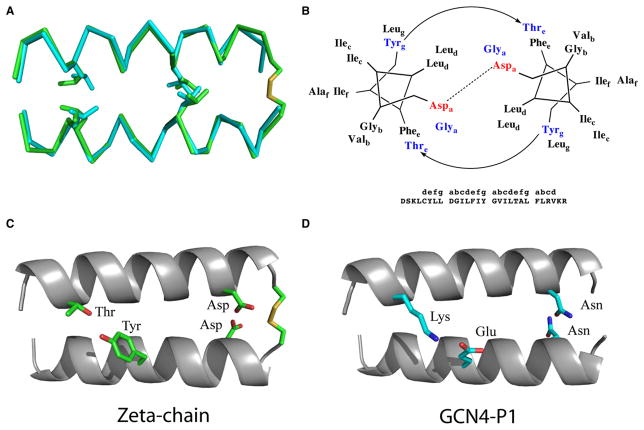
(A) Superposition of the TCR-ζ chain TM dimer (Call et al., 2006) and the GCN4 backbones (O’Shea et al., 1991). (B) Schematic of the crucial interactions within the TCR-ζ helical dimer. (C) TCR-ζ chain interactions including a Cys-disulfide, Asp-Asp hydrogen bonding, and Thr-Tyr hydrogen bonding. (D) The analogous GCN4-P1 interactions, including the Asn-Asn and Lys-Glu pairs.
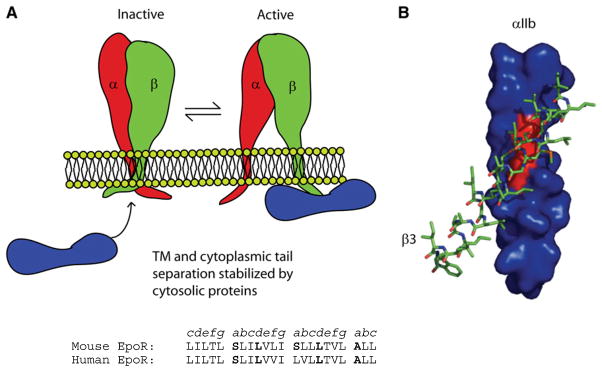
(A) Integrins are held inactive by interactions between α- and β-chain TM and soluble domains. Interactions between cytosolic proteins and integrin cytosolic tails stabilize TM separation and activation of the extracellular integrin ligand-binding site (Bennett, 2005). (B) A computational model of the β3-integrin TM domain packing against the GX3G motif of the αIIb-TM domain. Adapted from Li et al. (2005b).
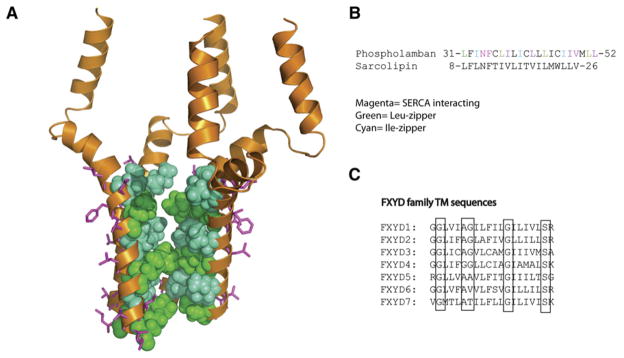
(A) An NMR structure of the phospholamban homopentamer (Oxenoid and Chou, 2005). In magenta are residues that mediate interactions with the SERCA channel. They are on the opposite face from the Leu- and Ile-zippers that mediate pentamerization (red and orange, respectively). (B) TM sequence of phospholamban showing important residues and the related TM sequence from sarcolipin. (C) Alignment of the FXYD protein family TM sequences, which regulate the Na/K-ATPase. Highly conserved and functionally important residues are highlighted. A mutation in the second conserved glycine, Gly41Arg, in the FXYD2 TM results in dominant negative primary hypomagnesemia due to aggregation of FXYD2 in the cytoplasm and misrouting of the Na,K-ATPase (Meij et al., 2000).
References
-
- Adamian L, Liang J. Interhelical hydrogen bonds and spatial motifs in membrane proteins: polar clamps and serine zippers. Proteins. 2002;47:209–218. - PubMed
-
- Afara MR, Trieber CA, Glaves JP, Young HS. Rational design of peptide inhibitors of the sarcoplasmic reticulum calcium pump. Biochemistry. 2006;45:8617–8627. - PubMed
-
- Akin C, Fumo G, Yavuz AS, Lipsky PE, Neckers L, Metcalfe DD. A novel form of mastocytosis associated with a transmembrane c-kit mutation and response to imatinib. Blood. 2004;103:3222–3225. - PubMed
-
- Bargmann CI, Hung MC, Weinberg RA. Multiple independent activations of the neu oncogene by a point mutation altering the transmembrane domain of p185. Cell. 1986;45:649–657. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
