

TAp73beta and DNp73beta activate the expression of the pro-survival caspase-2S
- PMID: 18611950
- PMCID: PMC2490756
- DOI: 10.1093/nar/gkn414
TAp73beta and DNp73beta activate the expression of the pro-survival caspase-2S
Abstract
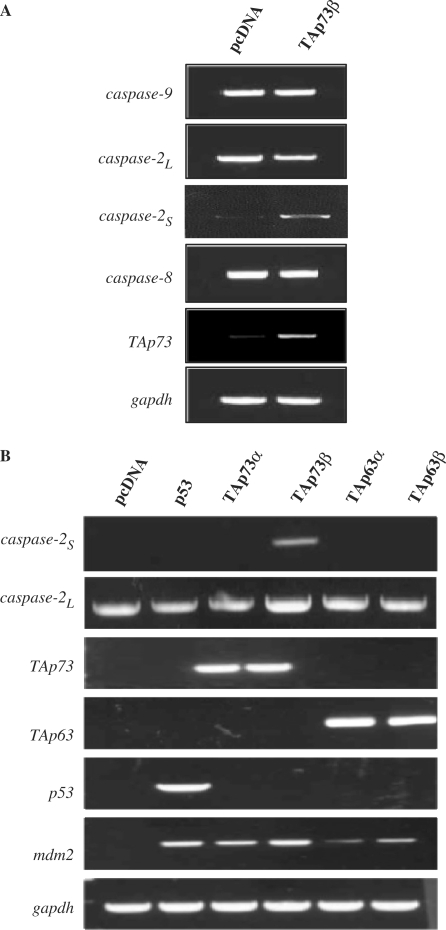
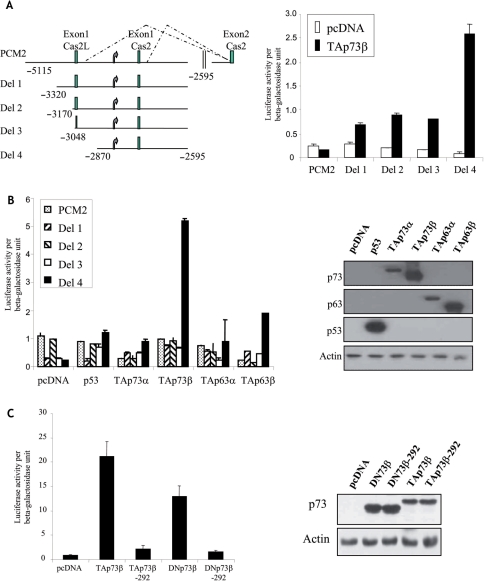
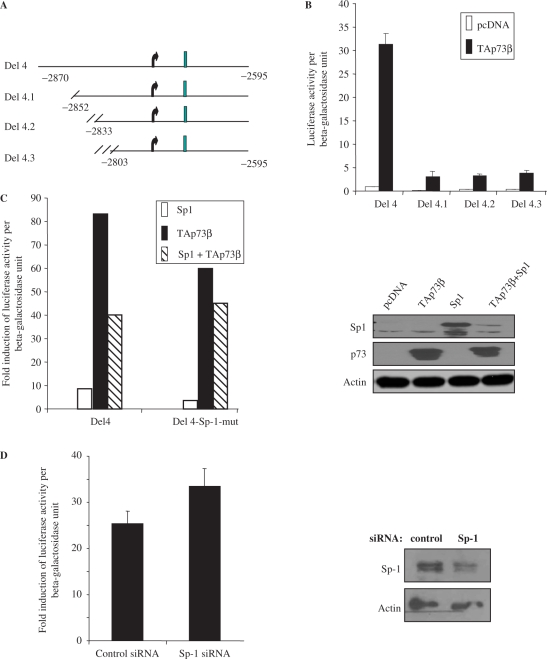
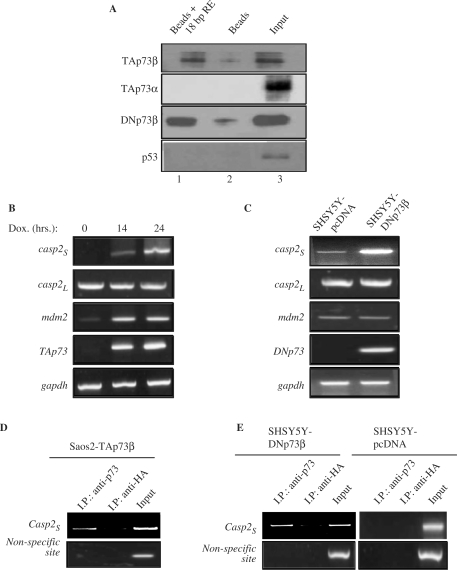
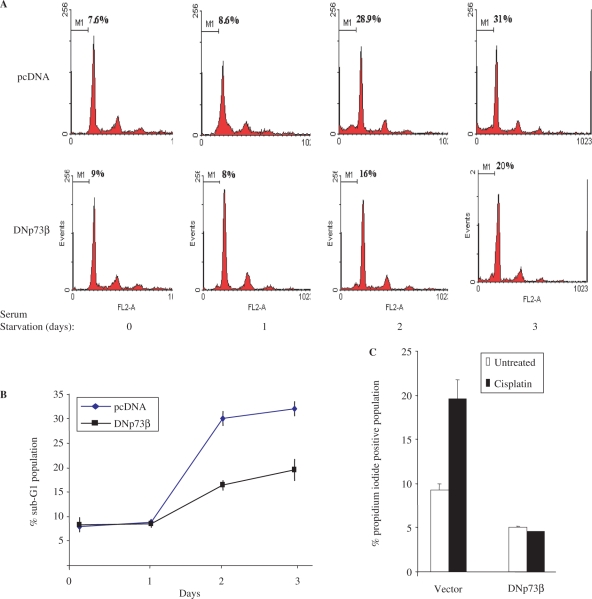
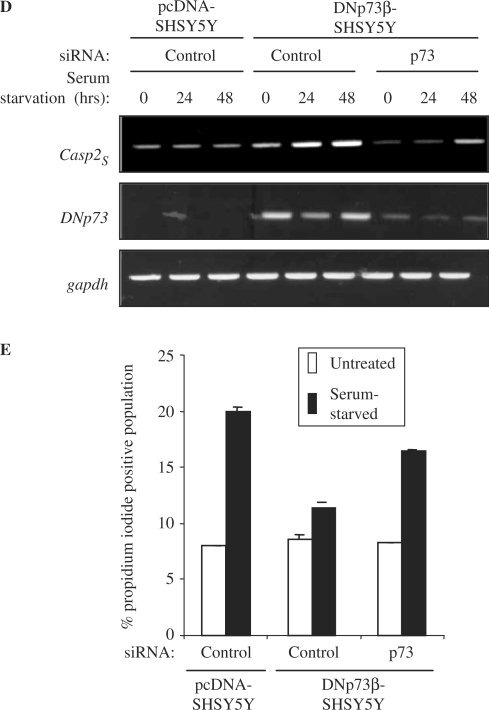
p73, the p53 homologue, exists as a transactivation-domain-proficient TAp73 or deficient deltaN(DN)p73 form. Expectedly, the oncogenic DNp73 that is capable of inactivating both TAp73 and p53 function, is over-expressed in cancers. However, the role of TAp73, which exhibits tumour-suppressive properties in gain or loss of function models, in human cancers where it is hyper-expressed is unclear. We demonstrate here that both TAp73 and DNp73 are able to specifically transactivate the expression of the anti-apoptotic member of the caspase family, caspase-2(S). Neither p53 nor TAp63 has this property, and only the p73beta form, but not the p73alpha form, has this competency. Caspase-2 promoter analysis revealed that a non-canonical, 18 bp GC-rich Sp-1-binding site-containing region is essential for p73beta-mediated activation. However, mutating the Sp-1-binding site or silencing Sp-1 expression did not affect p73beta's transactivation ability. In vitro DNA binding and in vivo chromatin immunoprecipitation assays indicated that p73beta is capable of directly binding to this region, and consistently, DNA binding p73 mutant was unable to transactivate caspase-2(S). Finally, DNp73beta over-expression in neuroblastoma cells led to resistance to cell death, and concomitantly to elevated levels of caspase-2(S.) Silencing p73 expression in these cells led to reduction of caspase-2(S) expression and increased cell death. Together, the data identifies caspase-2(S) as a novel transcriptional target common to both TAp73 and DNp73, and raises the possibility that TAp73 may be over-expressed in cancers to promote survival.
Figures
References
-
- Jost CA, Marin MC, Kaelin W.G., Jr p73 is a simian [correction of human] p53-related protein that can induce apoptosis. Nature. 1997;389:191–194. - PubMed
-
- Kaghad M, Bonnet H, Yang A, Creancier L, Biscan JC, Valent A, Minty A, Chalon P, Lelias JM, Dumont X, et al. Monoallelically expressed gene related to p53 at 1p36, a region frequently deleted in neuroblastoma and other human cancers. Cell. 1997;90:809–819. - PubMed
-
- Stiewe T, Zimmermann S, Frilling A, Esche H, Putzer BM. Transactivation-deficient DeltaTA-p73 acts as an oncogene. Cancer Res. 2002;62:3598–3602. - PubMed
-
- Stiewe T, Theseling CC, Putzer BM. Transactivation-deficient Delta TA-p73 inhibits p53 by direct competition for DNA binding: implications for tumorigenesis. J. Biol. Chem. 2002;277:14177–14185. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
