

Shaping cups into phagosomes and macropinosomes
- PMID: 18612320
- PMCID: PMC2851551
- DOI: 10.1038/nrm2447
Shaping cups into phagosomes and macropinosomes
Abstract
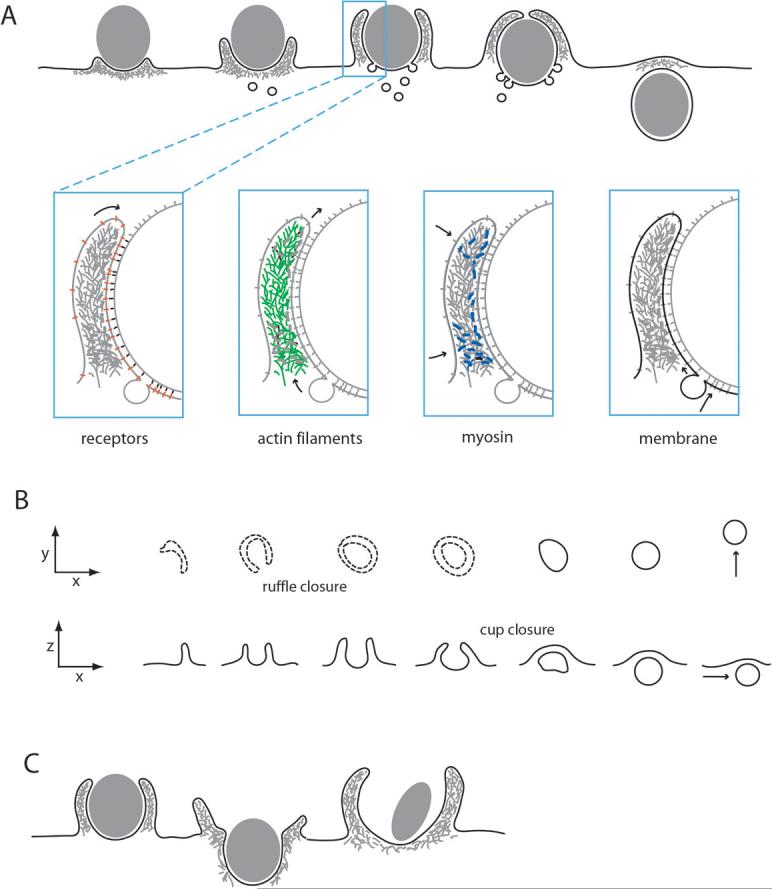
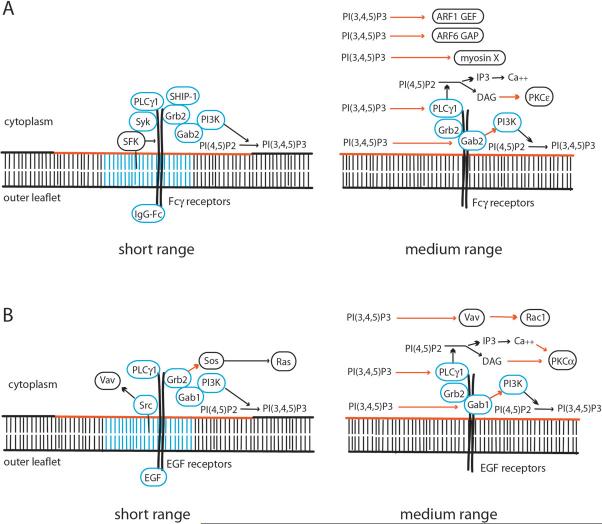
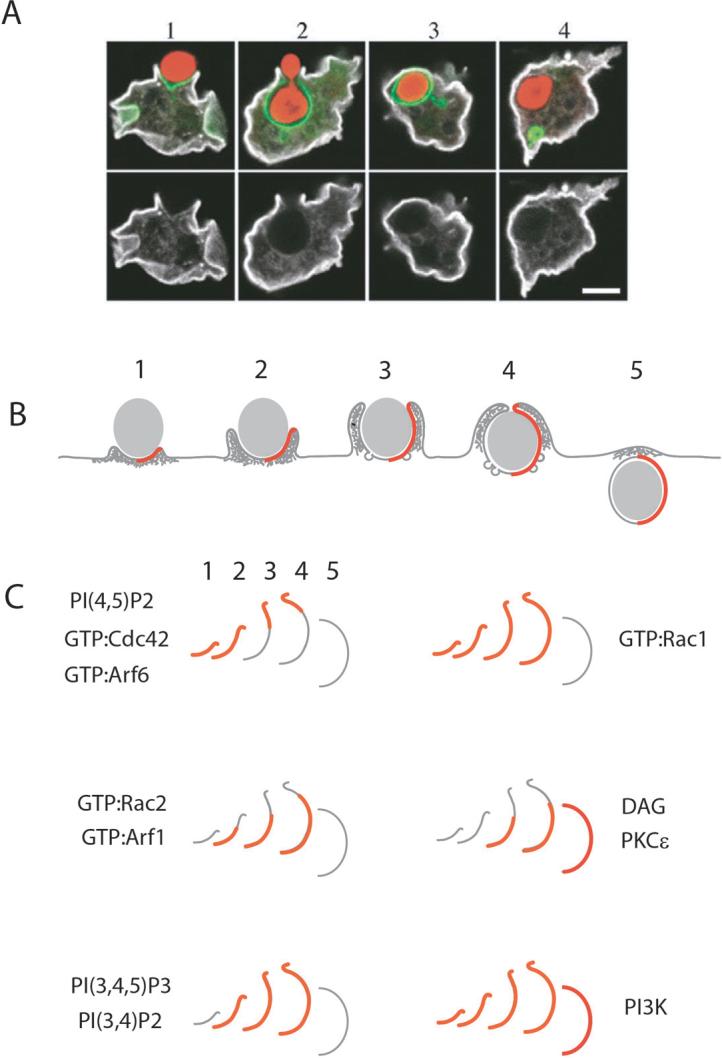
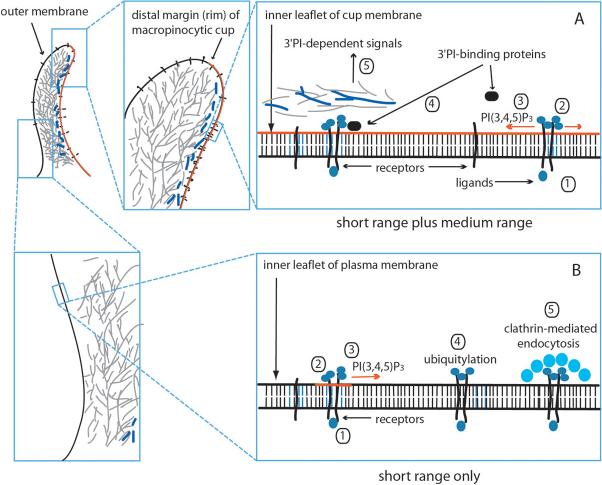
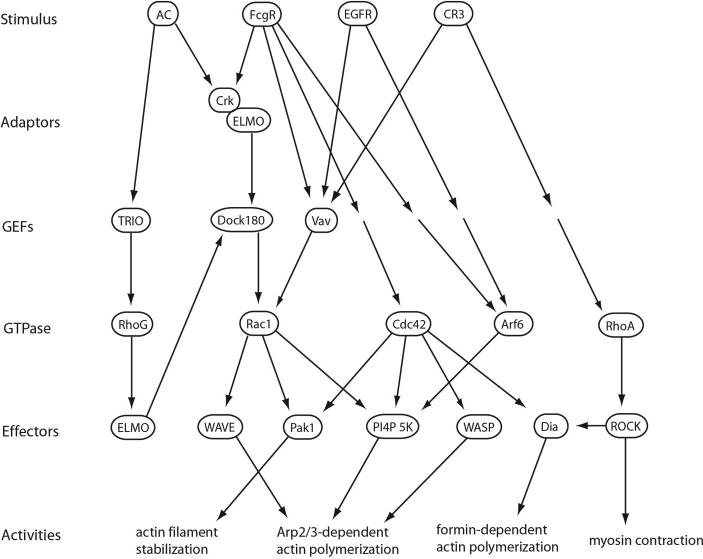
The ingestion of particles or cells by phagocytosis and of fluids by macropinocytosis requires the formation of large endocytic vacuolar compartments inside cells by the organized movements of membranes and the actin cytoskeleton. Fc-receptor-mediated phagocytosis is guided by the zipper-like progression of local, receptor-initiated responses that conform to particle geometry. By contrast, macropinosomes and some phagosomes form with little or no guidance from receptors. The common organizing structure is a cup-shaped invagination of the plasma membrane that becomes the phagosome or macropinosome. Recent studies, focusing on the physical properties of forming cups, indicate that a feedback mechanism regulates the signal transduction of phagocytosis and macropinocytosis.
Figures
References
-
- Stuart LM, Ezekowitz RA. Phagocytosis and comparative innate immunity: learning on the fly. Nat Rev Immunol. 2008;8:131–141. - PubMed
-
- Watts C, Amigorena S. Antigen traffic pathways in dendritic cells. Traffic. 2000;1:312–317. - PubMed
-
- Blander JM, Medzhitov R. On regulation of phagosome maturation and antigen presentation. Nat Immunol. 2006;7:1029–1035. - PubMed
-
- Reddien PW, Horvitz HR. The engulfment process of programmed cell death in caenorhabditis elegans. Annu Rev Cell Dev Biol. 2004;20:193–221. - PubMed
Highlighted References
-
- Park, et al. This paper identified a receptor phosphatidylserine presented on apoptotic cells and demonstrated a direct connection to Rac activation via ELMO and Dock180. 2007.
-
- Hall, et al. Demonstrated that the Rac GEF Vav was necessary for CR3-mediated phagocytosis but not for FcR-mediated phagocytosis. This finding is at odds with studies using other cells, which indicated a role for Vav-activated Rac in FcR but not CR3 phagocytosis. 2006.
-
- Amyere, et al. Demonstrated a role for PLCÎ downstream of PI3K during constitutive macropinocytosis in transformed cells. 2000.
-
- Botelho, et al. First demonstration of localized changes in phosphoinositides and diacylglycerol concentrations in unclosed phagocytic cups. 2000.
-
- Mercanti, et al. Demonstrated exclusion of membrane proteins from phagocytic and macropinocytic cups in Dictyostelium discoideum. 2006.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
