

LXR signaling couples sterol metabolism to proliferation in the acquired immune response
- PMID: 18614014
- PMCID: PMC2626438
- DOI: 10.1016/j.cell.2008.04.052
LXR signaling couples sterol metabolism to proliferation in the acquired immune response
Abstract
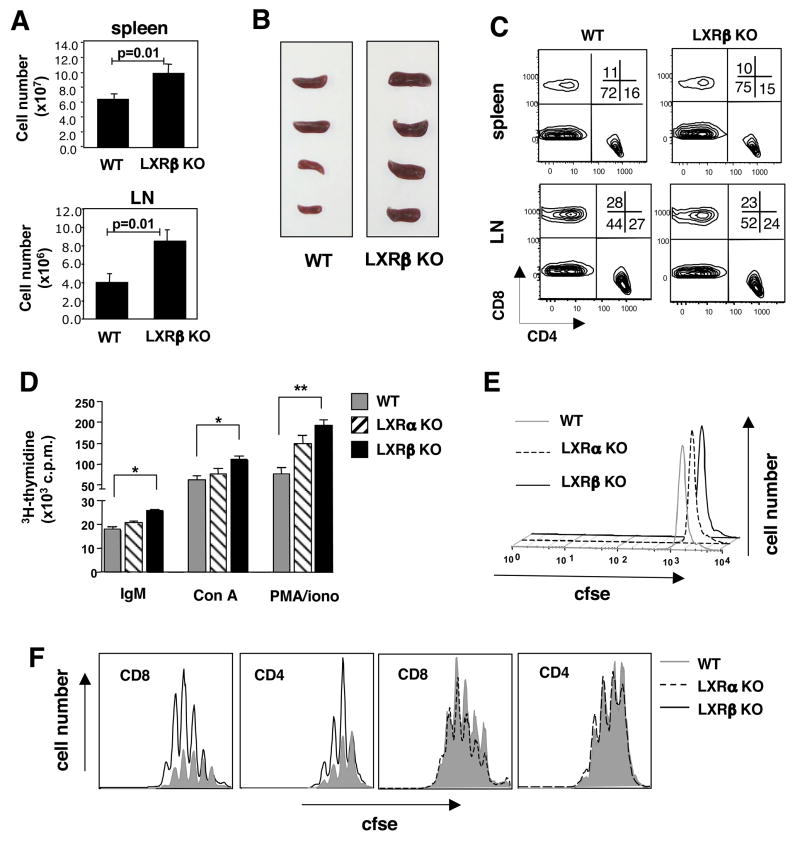
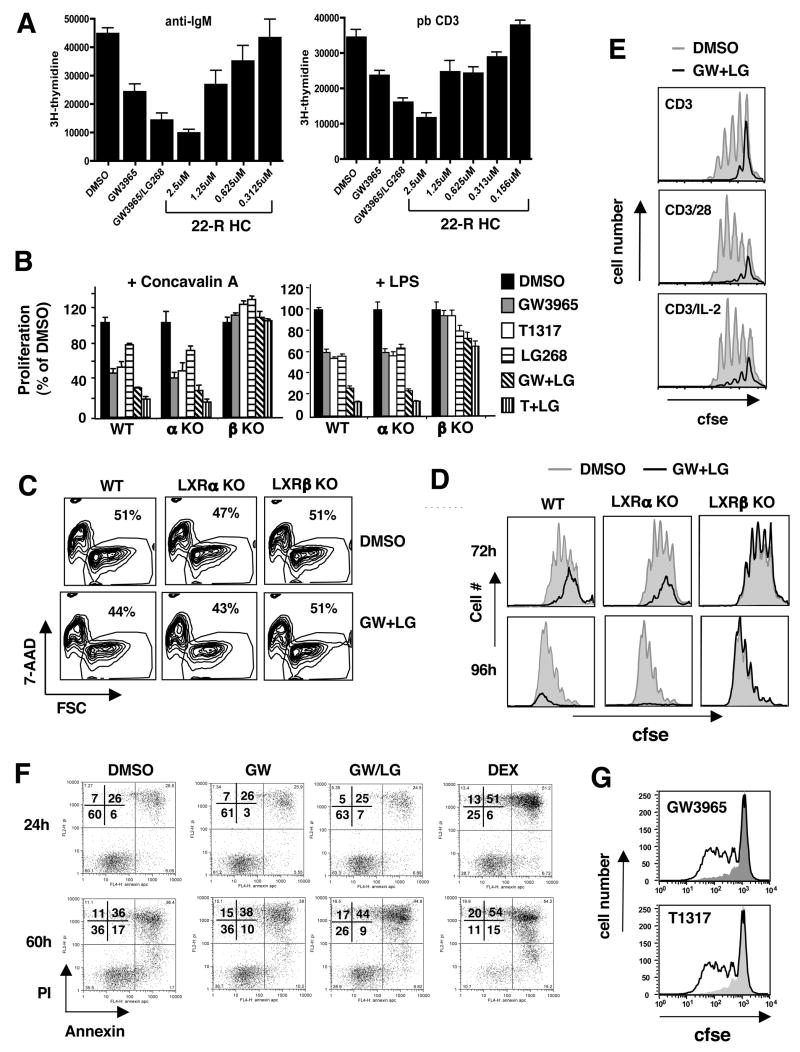
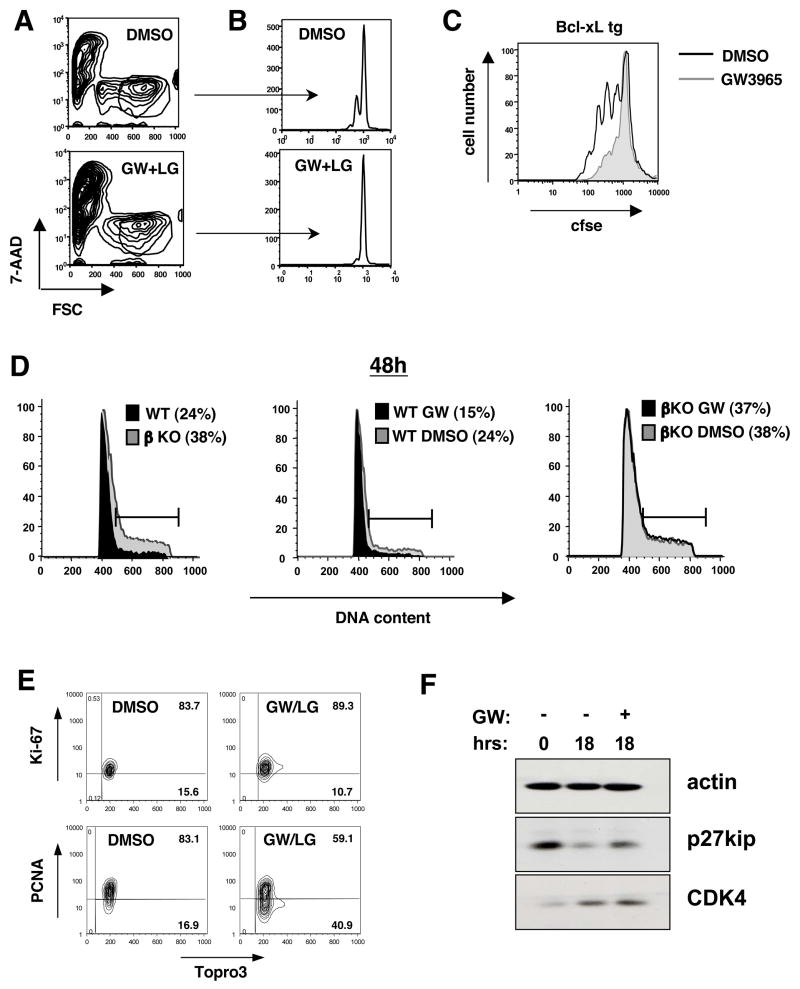
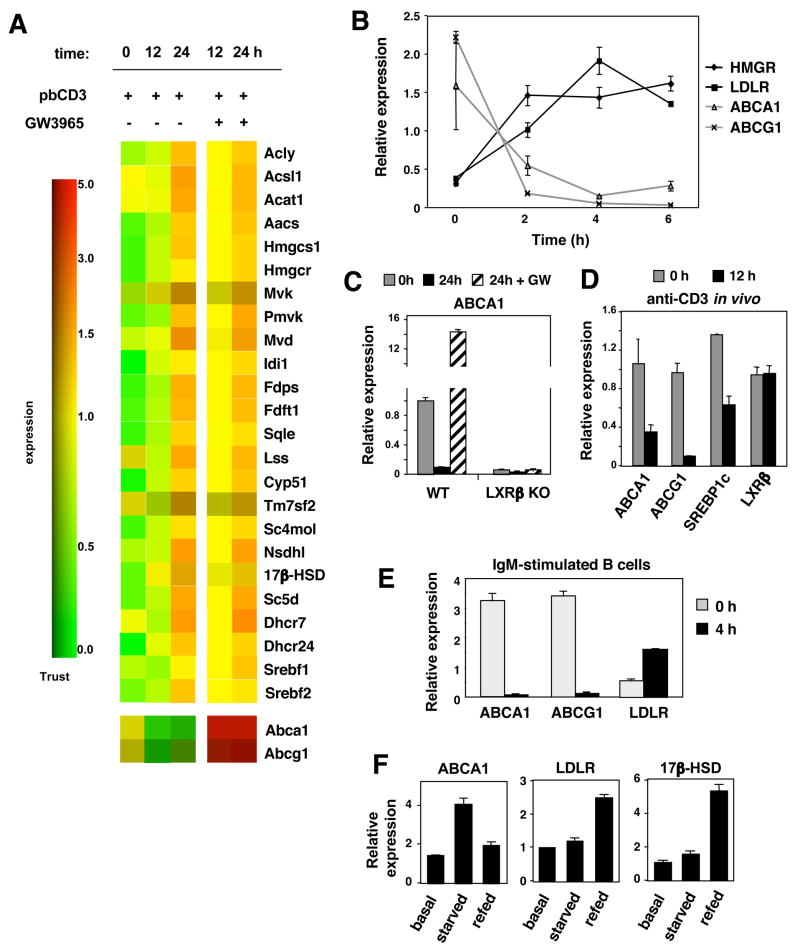
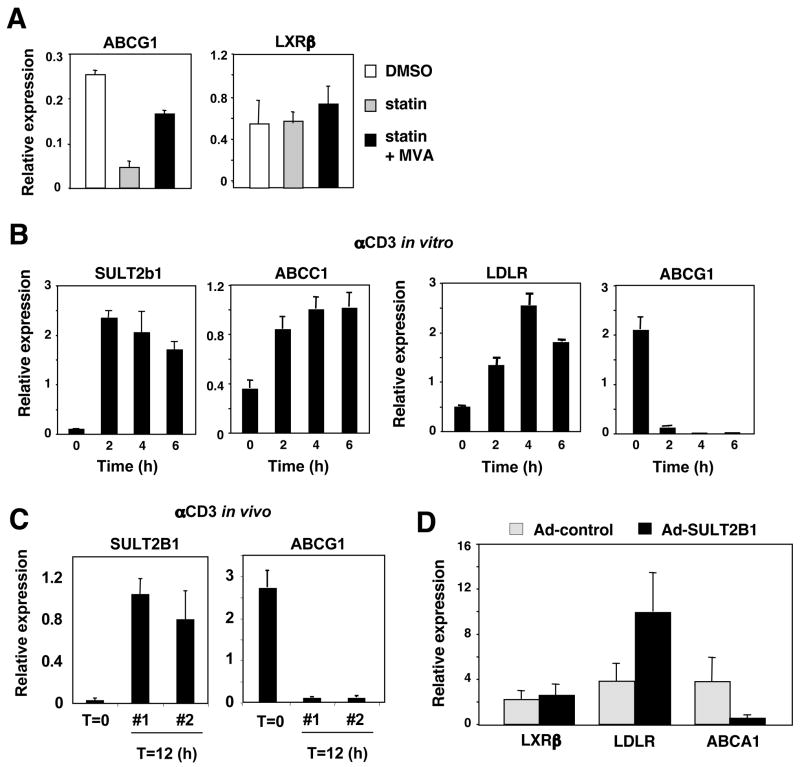
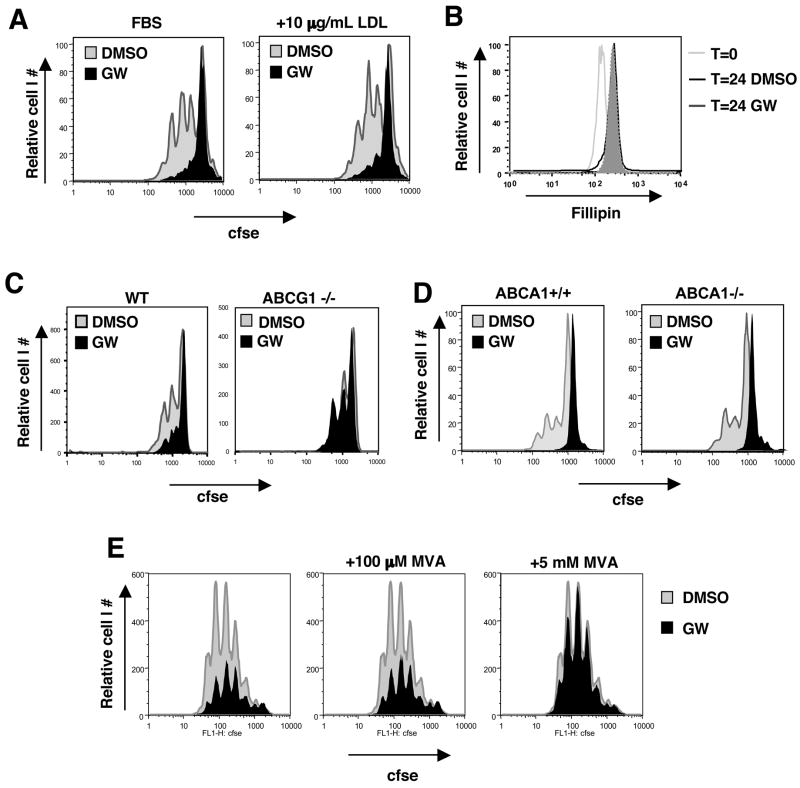
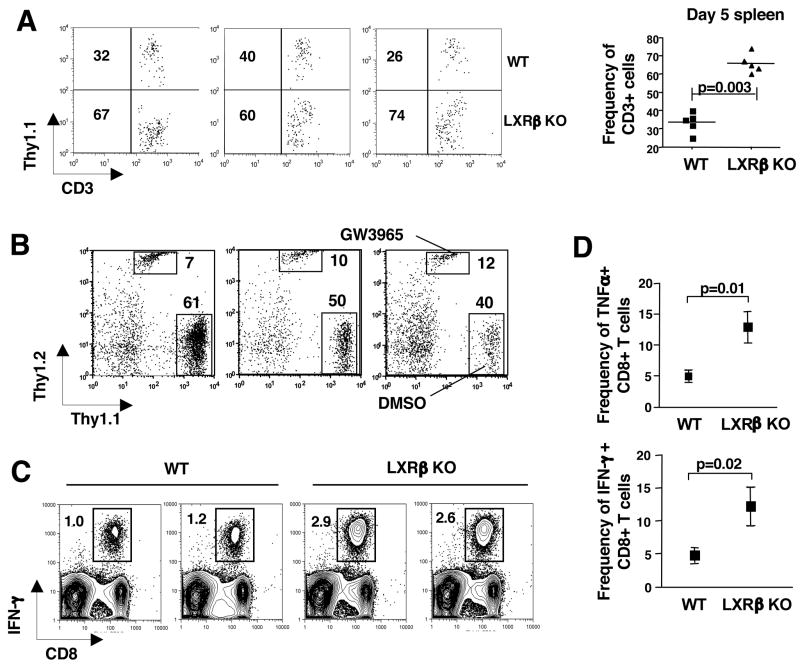
Cholesterol is essential for membrane synthesis; however, the mechanisms that link cellular lipid metabolism to proliferation are incompletely understood. We demonstrate here that cellular cholesterol levels in dividing T cells are maintained in part through reciprocal regulation of the LXR and SREBP transcriptional programs. T cell activation triggers induction of the oxysterol-metabolizing enzyme SULT2B1, consequent suppression of the LXR pathway for cholesterol transport, and promotion of the SREBP pathway for cholesterol synthesis. Ligation of LXR during T cell activation inhibits mitogen-driven expansion, whereas loss of LXRbeta confers a proliferative advantage. Inactivation of the sterol transporter ABCG1 uncouples LXR signaling from proliferation, directly linking sterol homeostasis to the antiproliferative action of LXR. Mice lacking LXRbeta exhibit lymphoid hyperplasia and enhanced responses to antigenic challenge, indicating that proper regulation of LXR-dependent sterol metabolism is important for immune responses. These results implicate LXR signaling in a metabolic checkpoint that modulates cell proliferation and immunity.
Figures
References
-
- Alves NL, Derks IA, Berk E, Spijker R, van Lier RA, Eldering E. The Noxa/Mcl-1 axis regulates susceptibility to apoptosis under glucose limitation in dividing T cells. Immunity. 2006;24:703–716. - PubMed
-
- Bromley SK, Burack WR, Johnson KG, Somersalo K, Sims TN, Sumen C, Davis MM, Shaw AS, Allen PM, Dustin ML. The immunological synapse. Annu Rev Immunol. 2001;19:375–396. - PubMed
-
- Brown MS, Goldstein JL. Suppression of 3-hydroxy-3-methylglutaryl coenzyme A reductase activity and inhibition of growth of human fibroblasts by 7-ketocholesterol. J Biol Chem. 1974;249:7306–7314. - PubMed
-
- Castrillo A, Joseph SB, Vaidya SA, Haberland M, Fogelman AM, Cheng G, Tontonoz P. Crosstalk between LXR and toll-like receptor signaling mediates bacterial and viral antagonism of cholesterol metabolism. Mol Cell. 2003;12:805–816. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
