

Chk2 suppresses the oncogenic potential of DNA replication-associated DNA damage
- PMID: 18614044
- PMCID: PMC2586815
- DOI: 10.1016/j.molcel.2008.04.028
Chk2 suppresses the oncogenic potential of DNA replication-associated DNA damage
Erratum in
- Mol Cell. 2008 Dec 26;32(6):889
Abstract
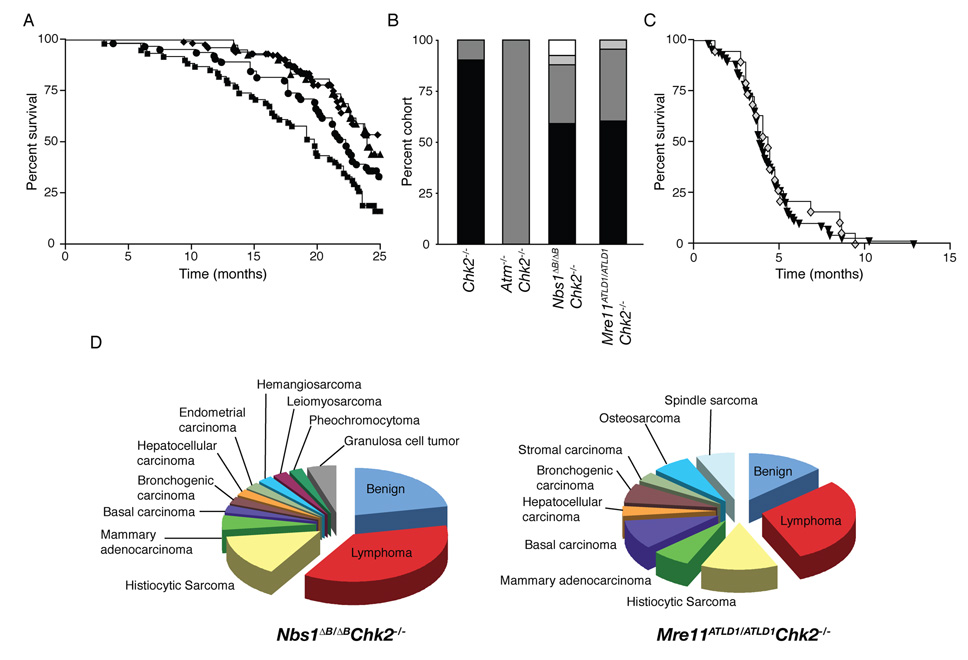
The Mre11 complex (Mre11, Rad50, and Nbs1) and Chk2 have been implicated in the DNA-damage response, an inducible process required for the suppression of malignancy. The Mre11 complex is predominantly required for repair and checkpoint activation in S phase, whereas Chk2 governs apoptosis. We examined the relationship between the Mre11 complex and Chk2 in the DNA-damage response via the establishment of Nbs1(DeltaB/DeltaB) Chk2(-/-) and Mre11(ATLD1/ATLD1) Chk2(-/-) mice. Chk2 deficiency did not modify the checkpoint defects or chromosomal instability of Mre11 complex mutants; however, the double-mutant mice exhibited synergistic defects in DNA-damage-induced p53 regulation and apoptosis. Nbs1(DeltaB/DeltaB) Chk2(-/-) and Mre11(ATLD1/ATLD1) Chk2(-/-) mice were also predisposed to tumors. In contrast, DNA-PKcs-deficient mice, in which G1-specific chromosome breaks are present, did not exhibit synergy with Chk2(-/-) mutants. These data suggest that Chk2 suppresses the oncogenic potential of DNA damage arising during S and G2 phases of the cell cycle.
Figures
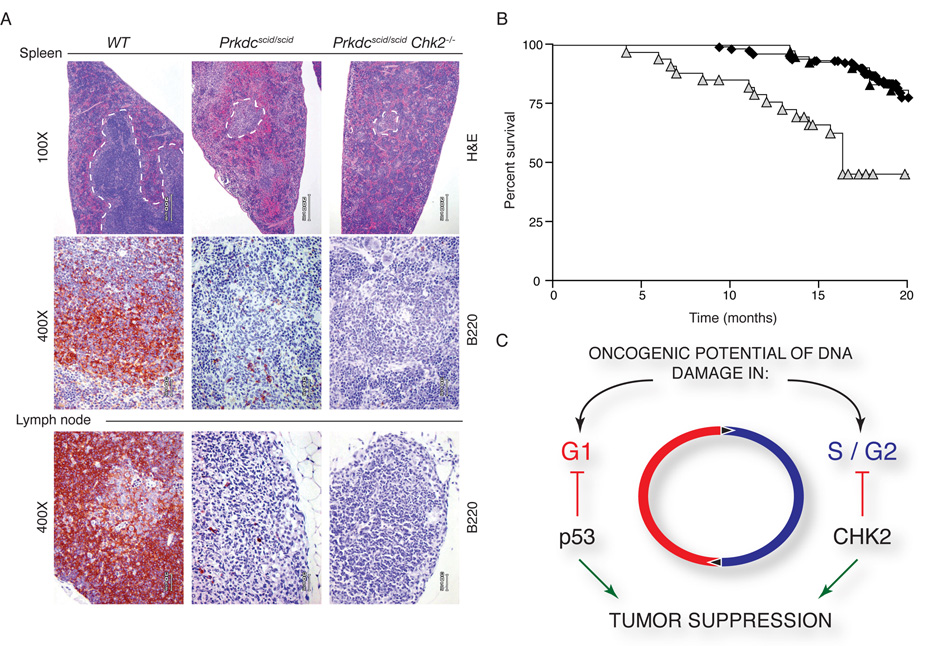
 , n=34) is plotted on a Kaplan Meier curve. Survival of Prkdcscid/scid Chk2−/− animals was decreased due to infections and related pathology but no tumors were evident. (C) Model for the role of Chk2 in tumor suppression. Chk2 is required for suppressing the oncogenic potential of DNA-replication associated breaks, such as those arising in Nbs1ΔB/ΔB or Mre11ATLD1ATLD1 mutants, but not for breaks in G0/G1 phase cells resulting from defects associated with the Prkdcscid/scid mutation.
, n=34) is plotted on a Kaplan Meier curve. Survival of Prkdcscid/scid Chk2−/− animals was decreased due to infections and related pathology but no tumors were evident. (C) Model for the role of Chk2 in tumor suppression. Chk2 is required for suppressing the oncogenic potential of DNA-replication associated breaks, such as those arising in Nbs1ΔB/ΔB or Mre11ATLD1ATLD1 mutants, but not for breaks in G0/G1 phase cells resulting from defects associated with the Prkdcscid/scid mutation.References
-
- Ahn JY, Schwarz JK, Piwnica-Worms H, Canman CE. Threonine 68 phosphorylation by ataxia telangiectasia mutated is required for efficient activation of Chk2 in response to ionizing radiation. Cancer Res. 2000;60:5934–5936. - PubMed
-
- Allen JB, Zhou Z, Siede W, Friedberg EC, Elledge SJ. The SAD1/RAD53 protein kinase controls multiple checkpoints and DNA damage-induced transcription in yeast. Genes & Dev. 1994;8:2401–2415. - PubMed
-
- Bahassi el M, Penner CG, Robbins SB, Tichy E, Feliciano E, Yin M, Liang L, Deng L, Tischfield JA, Stambrook PJ. The breast cancer susceptibility allele CHEK2* 1100delC promotes genomic instability in a knock-in mouse model. Mutat Res. 2007;616:201–209. - PubMed
-
- Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
