

Nonclassical mechanisms of progesterone action in the brain: II. Role of calmodulin-dependent protein kinase II in progesterone-mediated signaling in the hypothalamus of female rats
- PMID: 18617607
- PMCID: PMC2584600
- DOI: 10.1210/en.2008-0713
Nonclassical mechanisms of progesterone action in the brain: II. Role of calmodulin-dependent protein kinase II in progesterone-mediated signaling in the hypothalamus of female rats
Abstract
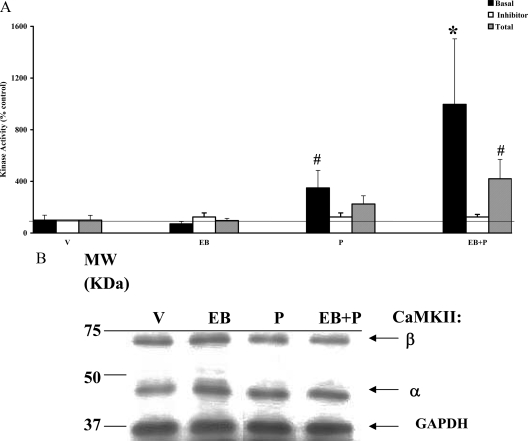
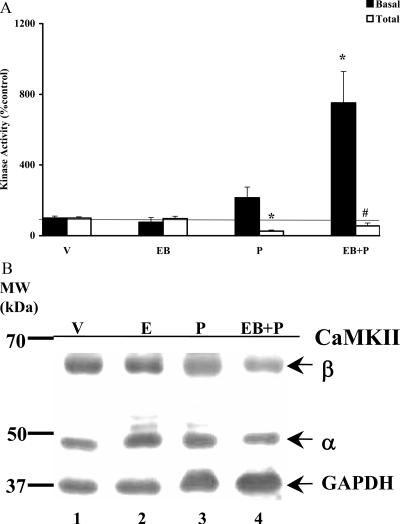
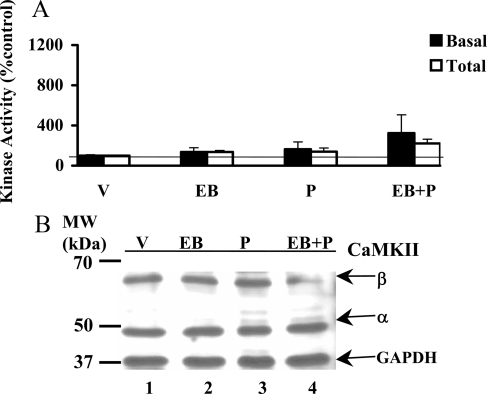
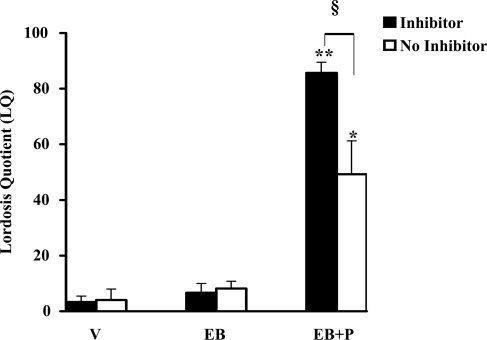
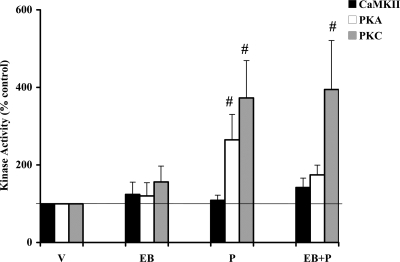
In addition to the activation of classical progestin receptor-dependent genomic pathway, progesterone (P) can activate nonclassical, membrane-initiated signaling pathways in the brain. We recently demonstrated rapid P activation of second-messenger kinases, protein kinase A, and protein kinase C in the ventromedial nucleus (VMN) and preoptic area (POA) of rat brain. To determine whether P can activate yet another Ca+2 dependent kinase, we examined the rapid P modulation of calcium and calmodulin-dependent protein kinase II (CaMKII) in the VMN and POA in female rats. A rapid P-initiated activation of CaMKII basal activity was observed in the VMN but not the POA at 30 min. Estradiol benzoate (EB) priming enhanced this CaMKII basal activity in both the VMN and POA. CaMKII protein levels and phosphorylation of Thr-286 moiety on CaMKII, however, remained unchanged with EB and/or P treatments, suggesting that the changes in the CaMKII kinase activity are due to rapid P modulation of the kinase activity and not its synthesis or autoactivation. Furthermore, intracerebroventricular (icv) administration of a CaMKII-specific inhibitor, KN-93, 30 min prior to the P infusion, in EB-primed, ovariectomized female rats inhibited CaMKII activation but not protein kinase A and protein kinase C activities. Interestingly, icv administration of KN-93 30 min prior to P infusion (icv) resulted in a reduction but not total inhibition of P-facilitated lordosis response in EB-primed female rats. These observations suggest a redundancy or, alternately, a hierarchy in the P-regulated activation of kinase signaling cascades in female reproductive behavior.
Figures
References
-
- Schumacher M, Coirini H, Pfaff DW, McEwen BS 1990 Behavioral effects of progesterone associated with rapid modulation of oxytocin receptors. Science 250:691–694 - PubMed
-
- Beyer C, Gonzalez-Mariscal G 1986 Elevation in hypothalamic cAMP as a common factor in the facilitation of lordosis in rodents: a working hypothesis. Ann NY Acad Sci 474:270–281 - PubMed
-
- Kow LM, Mobbs CV, Pfaff DW 1994 Role of second-messenger systems and neuronal activity in the regulation of lordosis by neurotransmitters neuropeptides and estrogen: a review. Neurosci Bio Behav Rev 18:251–268 - PubMed
-
- Pettiti N, Etgen AM 1989 Progesterone depression of norepinephrine-stimulated cAMP accumulation in hypothalamic slices. Brain Res Mol Brain Res 5:109–119 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
