

Osmotic regulation of Rab-mediated organelle docking
- PMID: 18619842
- PMCID: PMC2807628
- DOI: 10.1016/j.cub.2008.06.050
Osmotic regulation of Rab-mediated organelle docking
Abstract
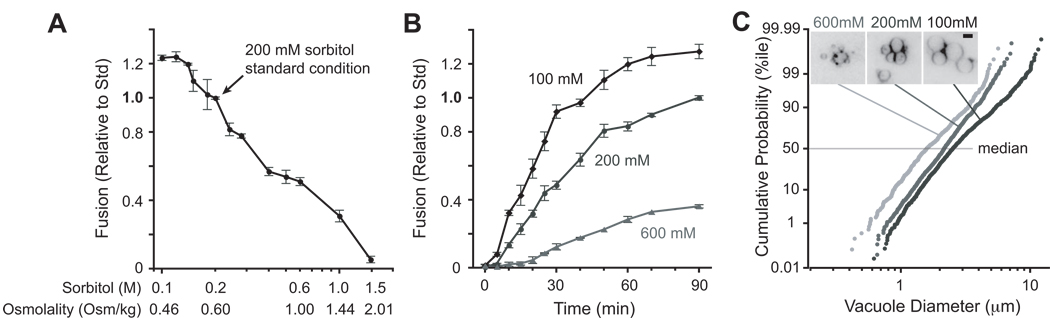
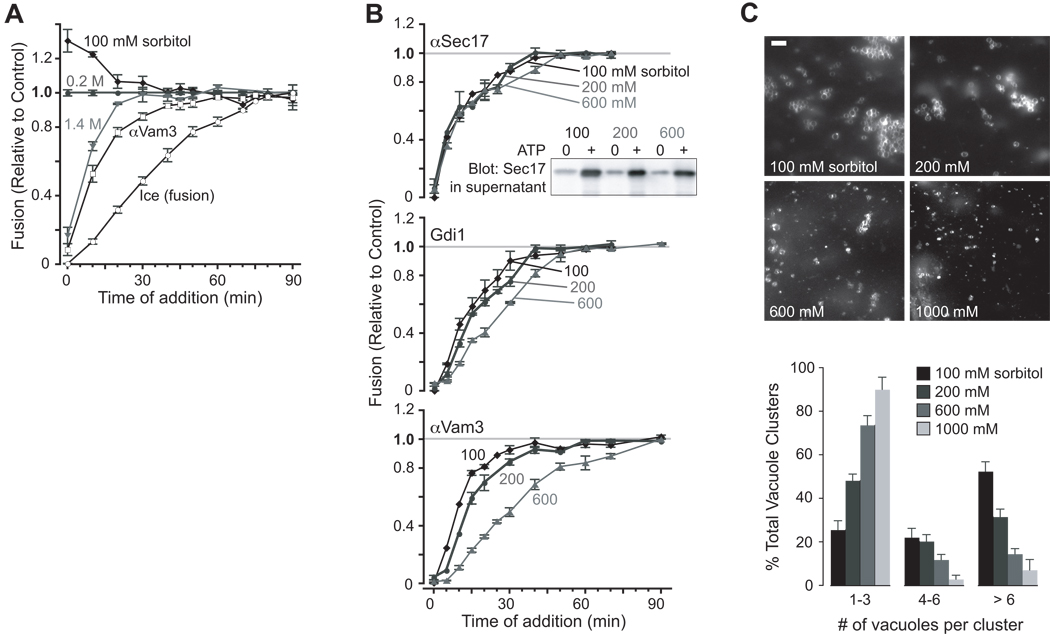
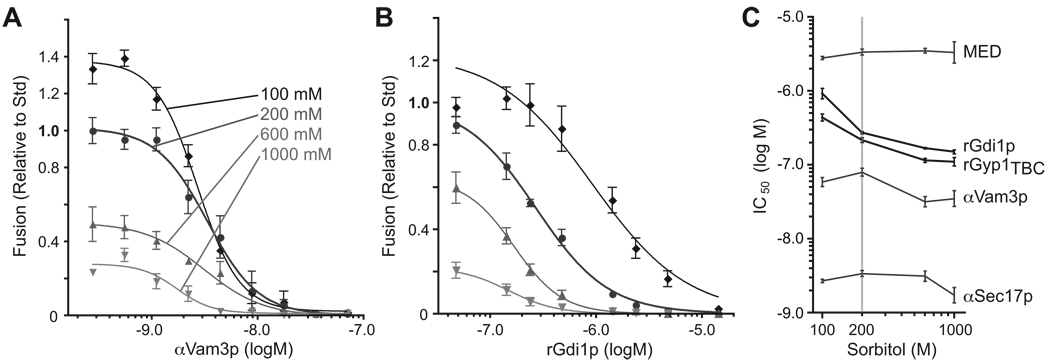
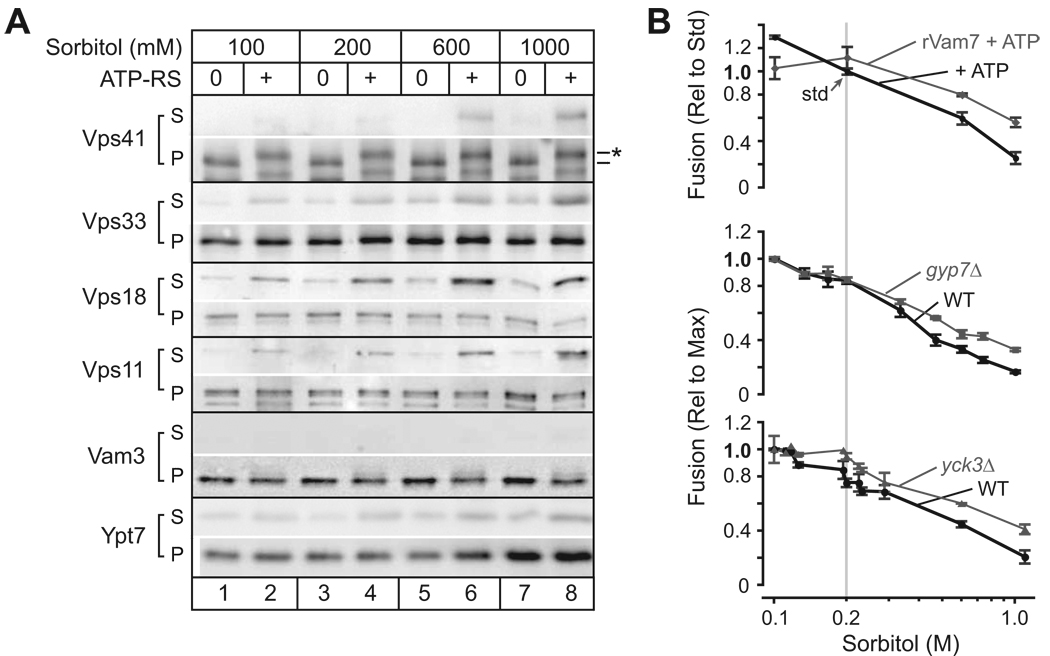
Osmotic gradients across organelle and plasma membranes modulate the rates of membrane fission and fusion; sufficiently large gradients can cause membrane rupture [1-6]. Hypotonic gradients applied to living yeast cells trigger prompt (within seconds) swelling and fusion of Saccharomyces cerevisiae vacuoles, whereas hypertonic gradients cause vacuoles to fragment on a slower time scale [7-11]. Here, we analyze the influence of osmotic strength on homotypic fusion of isolated yeast vacuoles. Consistent with previously reported in vivo results, we find that decreases in osmolyte concentration increase the rate and extent of vacuole fusion in vitro, whereas increases in osmolyte concentration prevent fusion. Unexpectedly, our results reveal that osmolytes regulate fusion by inhibiting early Rab-dependent docking or predocking events, not late events. Our experiments reveal an organelle-autonomous pathway that may control organelle surface-to-volume ratio, size, and copy number: Decreasing the osmolyte concentration in the cytoplasmic compartment accelerates Rab-mediated docking and fusion. By altering the relationship between the organelle surface and its enclosed volume, fusion in turn reduces the risk of membrane rupture.
Figures
References
-
- Tominaga T, Allen RD, Naitoh Y. Cyclic changes in the tension of the contractile vacuole complex membrane control its exocytotic cycle. J Exp Biol. 1998;201:2647–2658. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
