

Timing of sound-evoked potentials and spike responses in the inferior colliculus of awake bats
- PMID: 18621102
- PMCID: PMC2577224
- DOI: 10.1016/j.neuroscience.2008.06.031
Timing of sound-evoked potentials and spike responses in the inferior colliculus of awake bats
Abstract
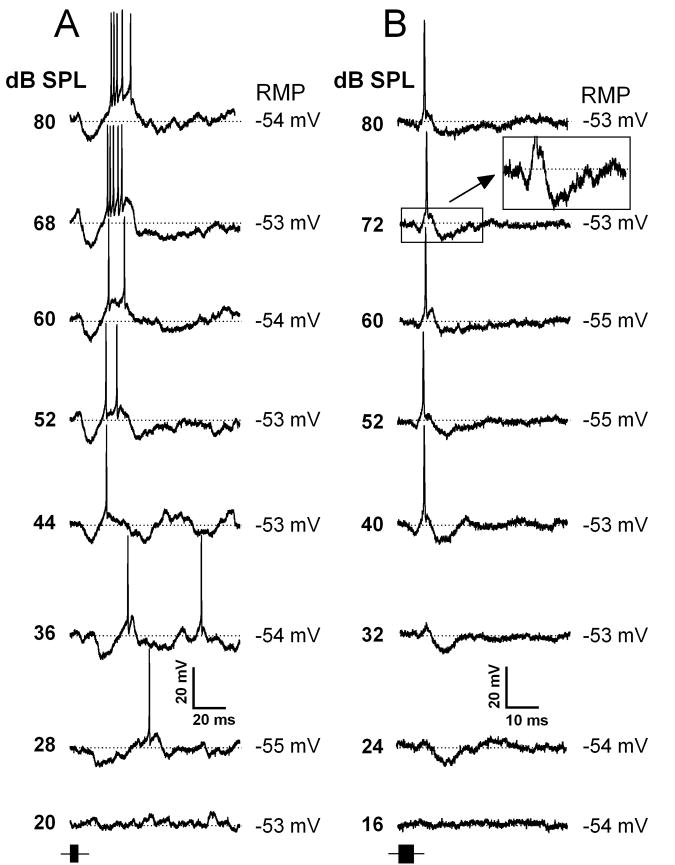
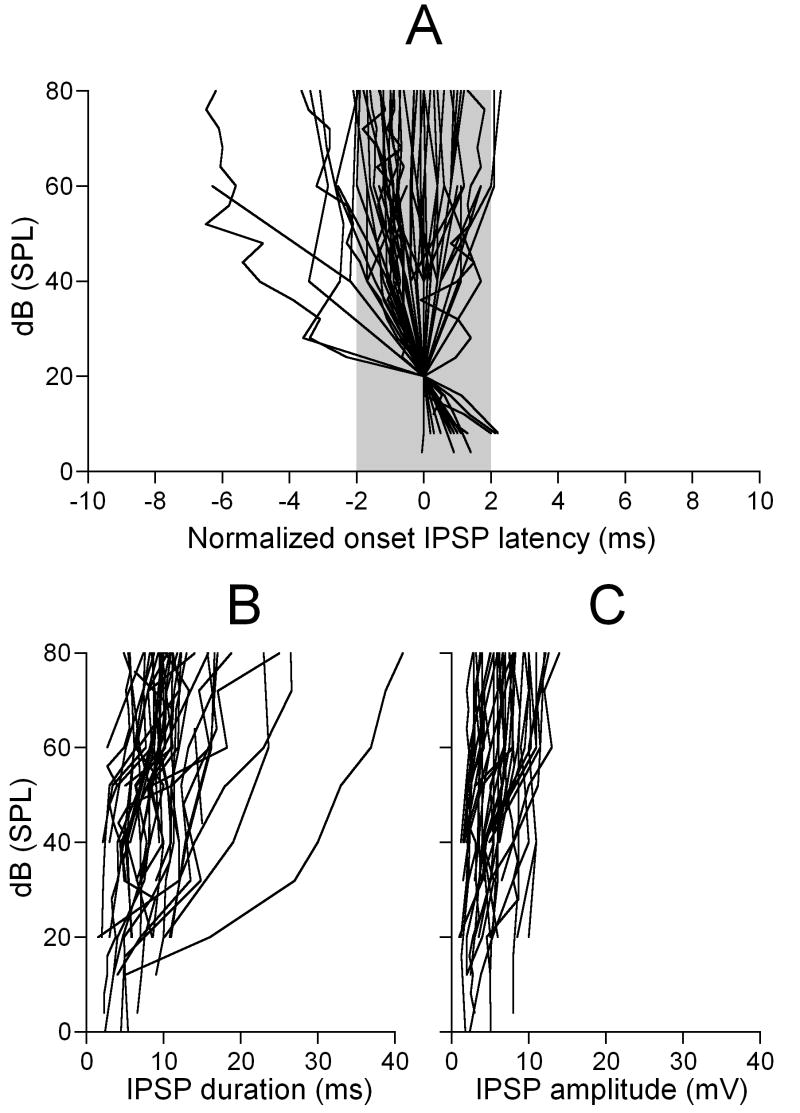
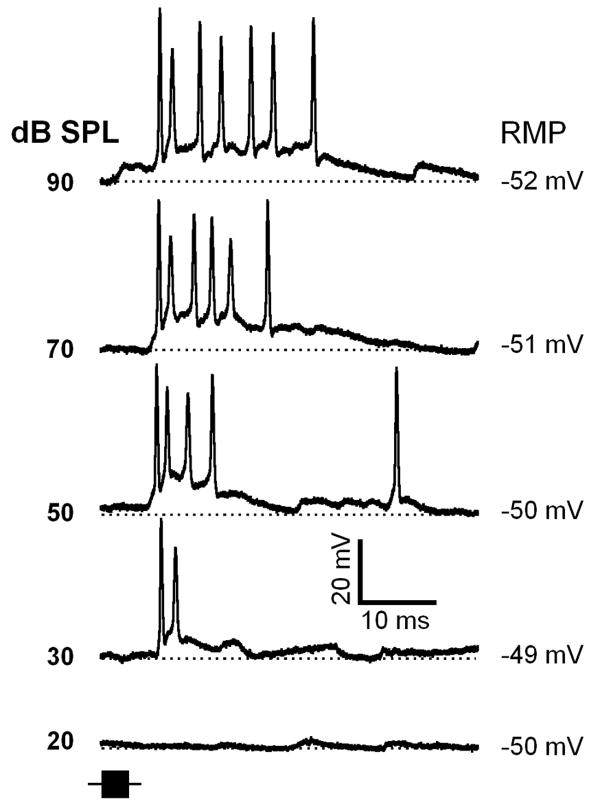
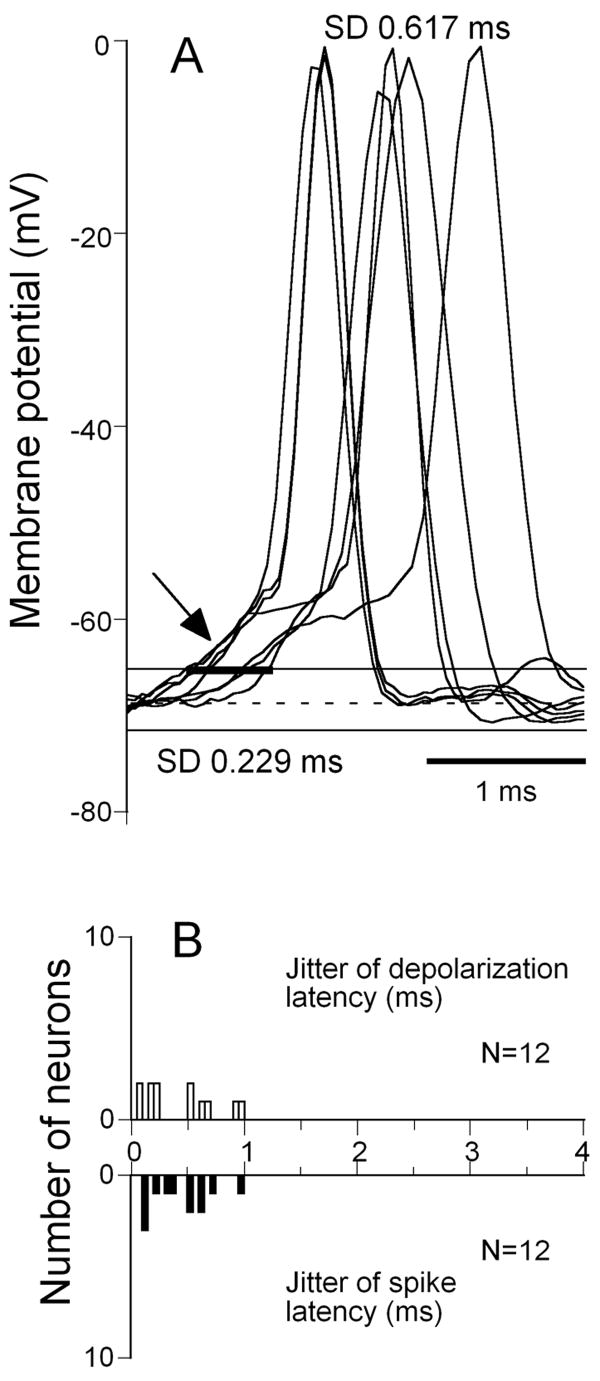
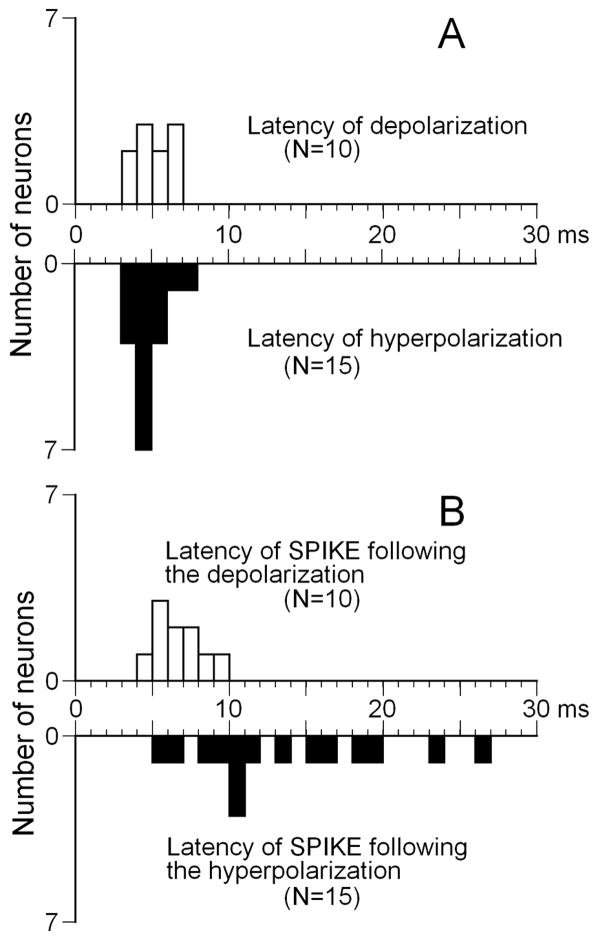
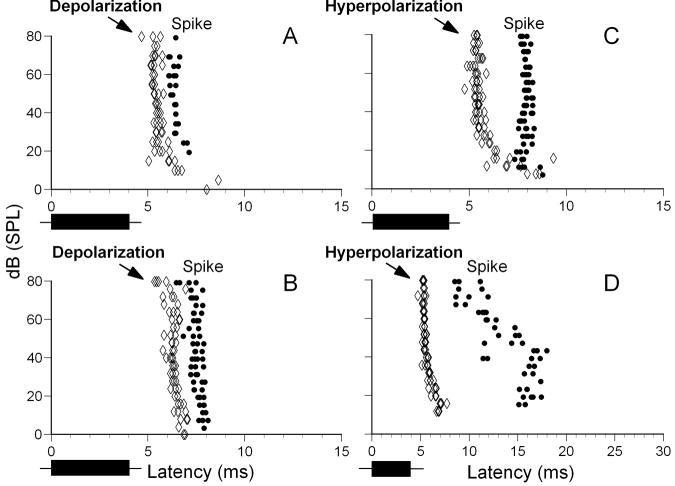
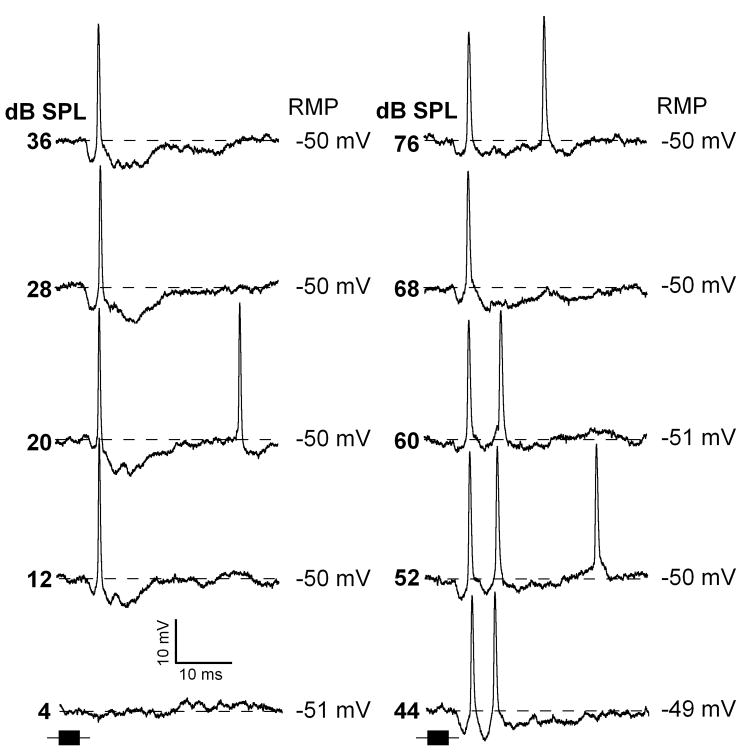
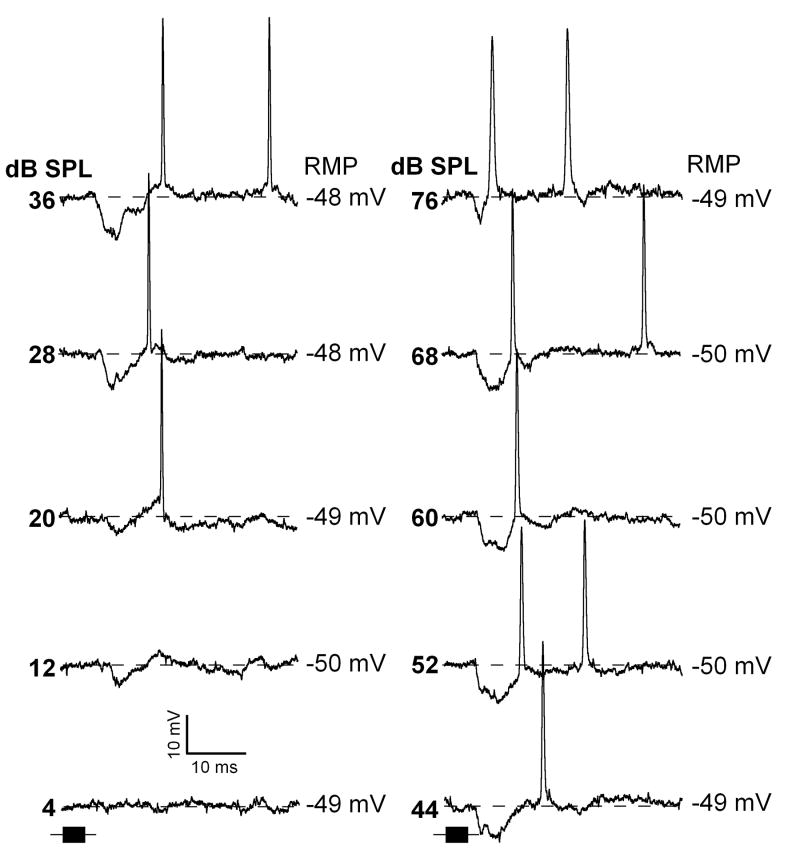
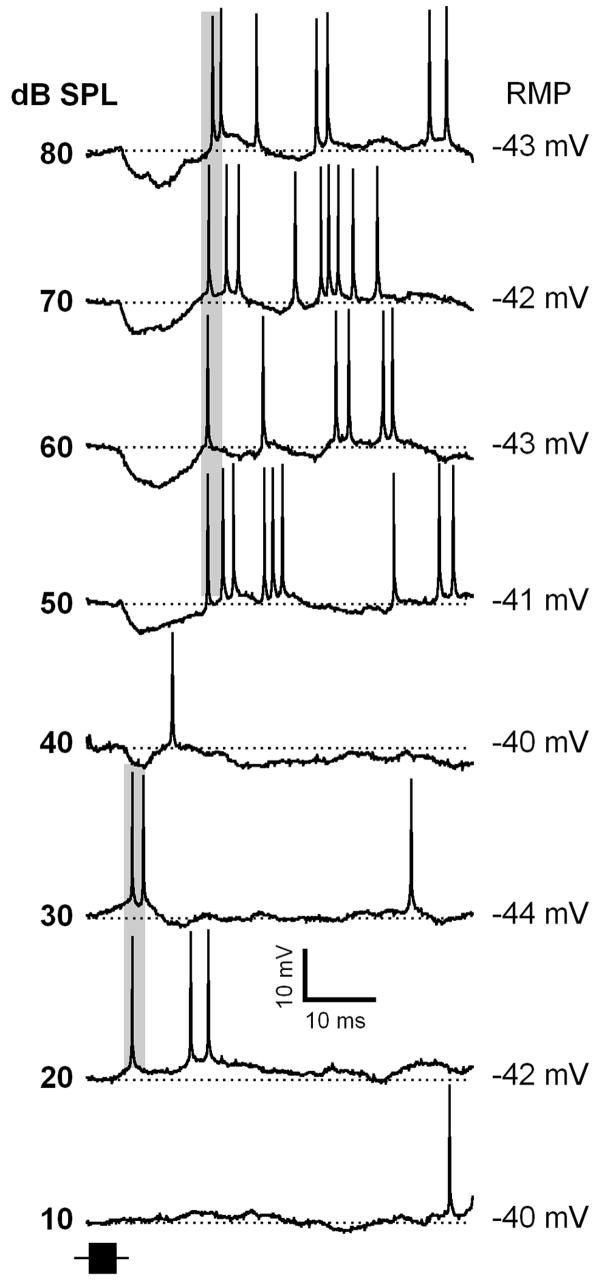
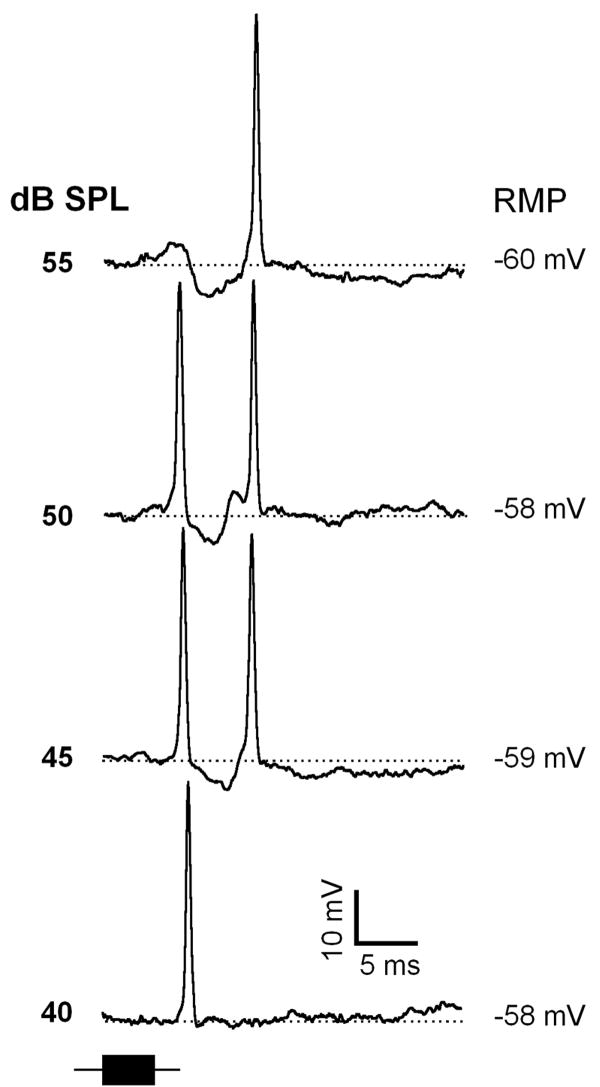
Neurons in the inferior colliculus (IC), one of the major integrative centers of the auditory system, process acoustic information converging from almost all nuclei of the auditory brain stem. During this integration, excitatory and inhibitory inputs arrive to auditory neurons at different time delays. Result of this integration determines timing of IC neuron firing. In the mammalian IC, the range of the first spike latencies is very large (5-50 ms). At present, a contribution of excitatory and inhibitory inputs in controlling neurons' firing in the IC is still under debate. In the present study we assess the role of excitation and inhibition in determining first spike response latency in the IC. Postsynaptic responses were recorded to pure tones presented at neuron's characteristic frequency or to downward frequency modulated sweeps in awake bats. There are three main results emerging from the present study: (1) the most common response pattern in the IC is hyperpolarization followed by depolarization followed by hyperpolarization, (2) latencies of depolarizing or hyperpolarizing responses to tonal stimuli are short (3-7 ms) whereas the first spike latencies may vary to a great extent (4-26 ms) from one neuron to another, and (3) high threshold hyperpolarization preceded long latency spikes in IC neurons exhibiting paradoxical latency shift. Our data also show that the onset hyperpolarizing potentials in the IC have very small jitter (<100 micros) across repeated stimulus presentations. The results of this study suggest that inhibition, arriving earlier than excitation, may play a role as a mechanism for delaying the first spike latency in IC neurons.
Figures

References
-
- Aitkin LM, Anderson DJ, Brugge JF. Tonotopic organization and discharge characteristics of single neurons in nuclei of the lateral lemniscus of the cat. J Neurophysiol. 1970;33:421–440. - PubMed
-
- Berkowitz A, Suga N. Neural mechanisms of ranging are different in two species of bats. Hear Res. 1989;41:255–264. - PubMed
-
- Binns KE. The synaptic pharmacology underlying sensory processing in the superior colliculus. Prog Neurobiol. 1999;59:129–159. - PubMed
-
- Blitz DM, Regehr WG. Retinogeniculate synaptic properties controlling spike number and timing in relay neuron. J Neurophysiol. 2003;90:2438–2450. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
