

In silico identified CCR4 antagonists target regulatory T cells and exert adjuvant activity in vaccination
- PMID: 18621704
- PMCID: PMC2481334
- DOI: 10.1073/pnas.0803453105
In silico identified CCR4 antagonists target regulatory T cells and exert adjuvant activity in vaccination
Abstract
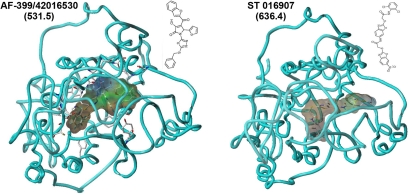
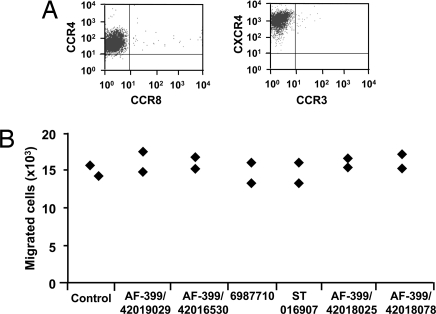
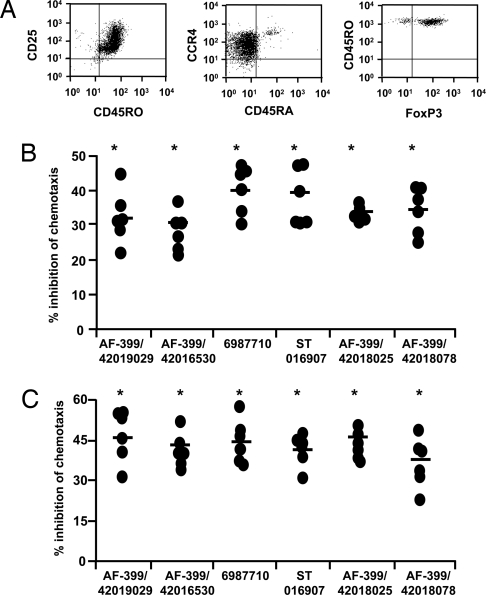
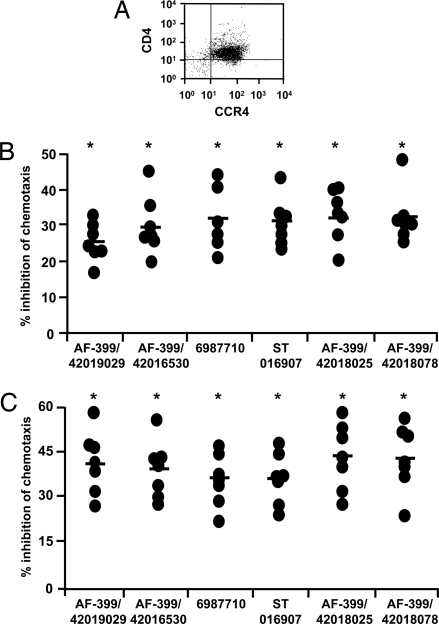
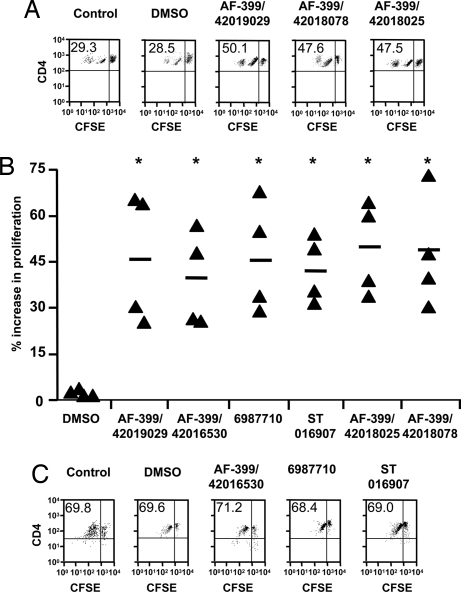
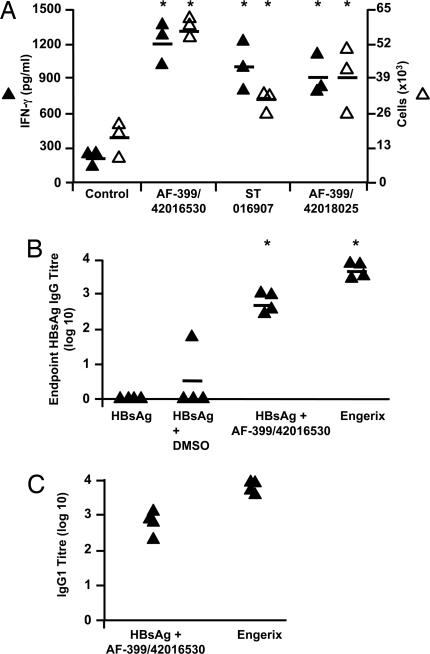
Adjuvants are substances that enhance immune responses and thus improve the efficacy of vaccination. Few adjuvants are available for use in humans, and the one that is most commonly used (alum) often induces suboptimal immunity for protection against many pathogens. There is thus an obvious need to develop new and improved adjuvants. We have therefore taken an approach to adjuvant discovery that uses in silico modeling and structure-based drug-design. As proof-of-principle we chose to target the interaction of the chemokines CCL22 and CCL17 with their receptor CCR4. CCR4 was posited as an adjuvant target based on its expression on CD4(+)CD25(+) regulatory T cells (Tregs), which negatively regulate immune responses induced by dendritic cells (DC), whereas CCL17 and CCL22 are chemotactic agents produced by DC, which are crucial in promoting contact between DC and CCR4(+) T cells. Molecules identified by virtual screening and molecular docking as CCR4 antagonists were able to block CCL22- and CCL17-mediated recruitment of human Tregs and Th2 cells. Furthermore, CCR4 antagonists enhanced DC-mediated human CD4(+) T cell proliferation in an in vitro immune response model and amplified cellular and humoral immune responses in vivo in experimental models when injected in combination with either Modified Vaccinia Ankara expressing Ag85A from Mycobacterium tuberculosis (MVA85A) or recombinant hepatitis B virus surface antigen (rHBsAg) vaccines. The significant adjuvant activity observed provides good evidence supporting our hypothesis that CCR4 is a viable target for rational adjuvant design.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Guy B. The perfect mix: Recent progress in adjuvant research. Nat Rev Microbiol. 2007;5:505–517. - PubMed
-
- Schijns VE, Degen WG. Vaccine immunopotentiators of the future. Clin Pharmacol Ther. 2007;82:750–755. - PubMed
-
- Davies MN, Flower DR. Harnessing bioinformatics to discover new vaccines. Drug Discov Today. 2007;12:389–395. - PubMed
-
- Miyara M, Sakaguchi S. Natural regulatory T cells: Mechanisms of suppression. Trends Mol Med. 2007;13:108–116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
