

Monovalent and divalent promoted GAAA tetraloop-receptor tertiary interactions from freely diffusing single-molecule studies
- PMID: 18621836
- PMCID: PMC2553128
- DOI: 10.1529/biophysj.108.134346
Monovalent and divalent promoted GAAA tetraloop-receptor tertiary interactions from freely diffusing single-molecule studies
Abstract
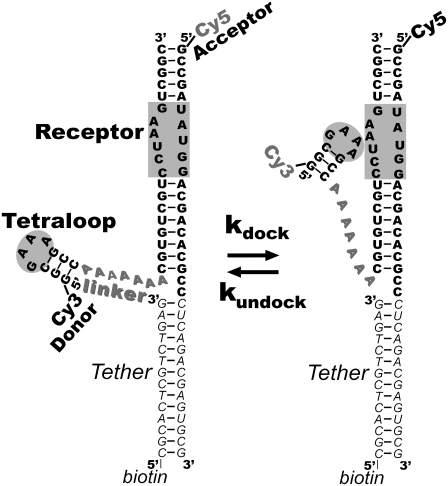
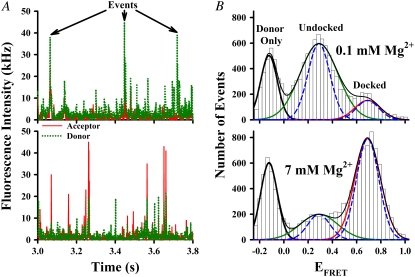
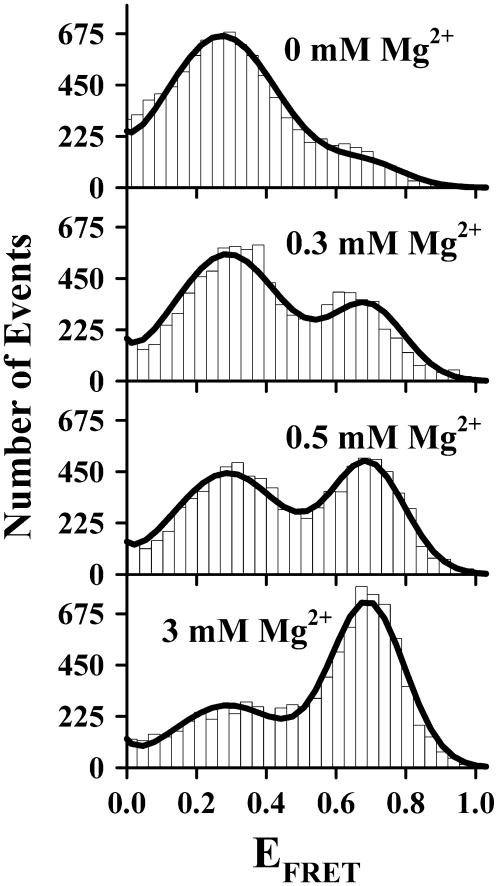
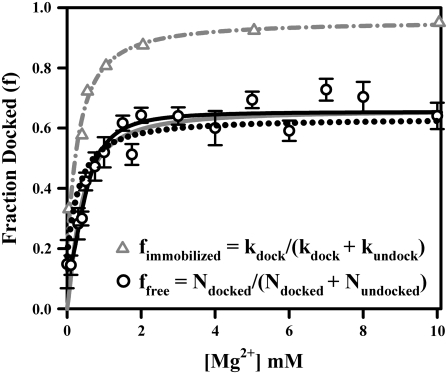
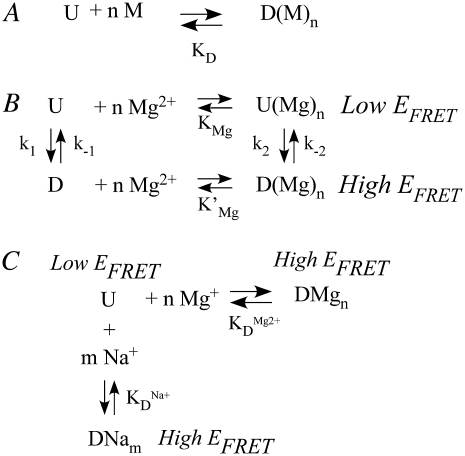
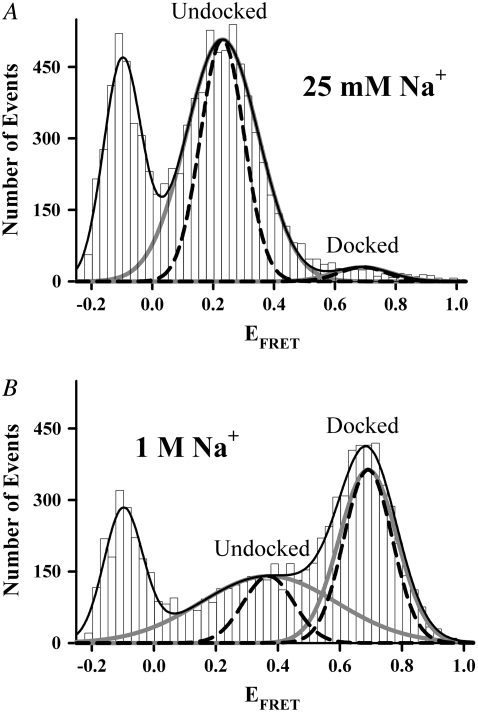
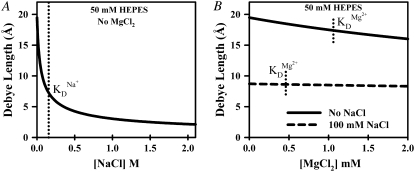
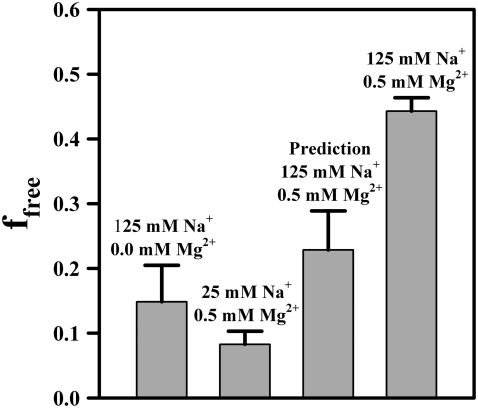
Proper assembly of RNA into catalytically active three-dimensional structures requires multiple tertiary binding interactions, individual characterization of which is crucial to a detailed understanding of global RNA folding. This work focuses on single-molecule fluorescence studies of freely diffusing RNA constructs that isolate the GAAA tetraloop-receptor tertiary interaction. Freely diffusing conformational dynamics are explored as a function of Mg(2+) and Na(+) concentration, both of which promote facile docking, but with 500-fold different affinities. Systematic shifts in mean fluorescence resonance energy transfer efficiency values and line widths with increasing [Na(+)] are observed for the undocked species and can be interpreted with a Debye model in terms of electrostatic relaxation and increased flexibility in the RNA. Furthermore, we identify a 34 +/- 2% fraction of freely diffusing RNA constructs remaining undocked even at saturating [Mg(2+)] levels, which agrees quantitatively with the 32 +/- 1% fraction previously reported for immobilized constructs. This verifies that the kinetic heterogeneity observed in the docking rates is not the result of surface tethering. Finally, the K(D) value and Hill coefficient for [Mg(2+)]-dependent docking decrease significantly for [Na(+)] = 25 mM vs. 125 mM, indicating Mg(2+) and Na(+) synergy in the RNA folding process.
Figures


References
-
- Gesteland, R. F., T. R. Cech, and J. F. Atkins. 2006. The RNA World: The Nature of Modern RNA Suggests a Prebiotic RNA. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Tinoco, I., and C. Bustamante. 1999. How RNA folds. J. Mol. Biol. 293:271–281. - PubMed
-
- Batey, R. T., R. P. Rambo, and J. A. Doudna. 1999. Tertiary motifs in RNA structure and folding. Angew. Chem. Int. Ed. 38:2327–2343. - PubMed
-
- Young, B. T., and S. K. Silverman. 2002. The GAAA tetraloop-receptor interaction contributes differentially to folding thermodynamics and kinetics for the P4-P6 RNA domain. Biochemistry. 41:12271–12276. - PubMed
-
- Bartley, L. E., X. W. Zhuang, R. Das, S. Chu, and D. Herschlag. 2003. Exploration of the transition state for tertiary structure formation between an RNA helix and a large structured RNA. J. Mol. Biol. 328:1011–1026. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
