

Mutation of the plastidial alpha-glucan phosphorylase gene in rice affects the synthesis and structure of starch in the endosperm
- PMID: 18621947
- PMCID: PMC2518224
- DOI: 10.1105/tpc.107.054007
Mutation of the plastidial alpha-glucan phosphorylase gene in rice affects the synthesis and structure of starch in the endosperm
Abstract
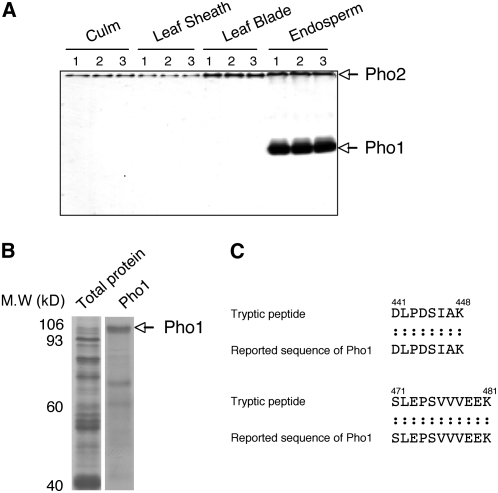
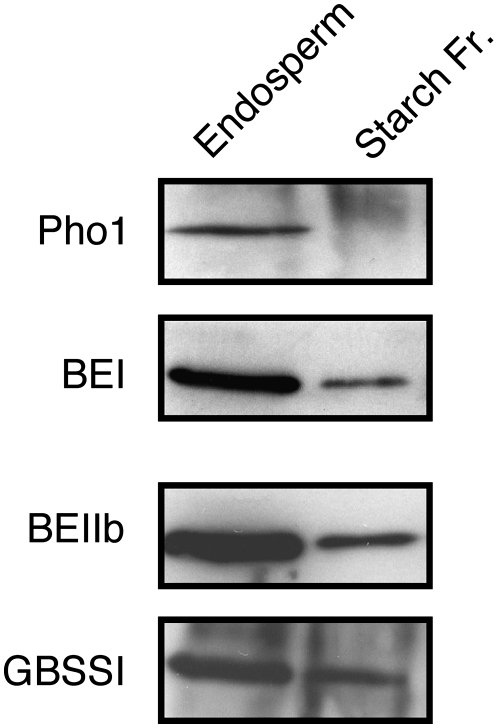
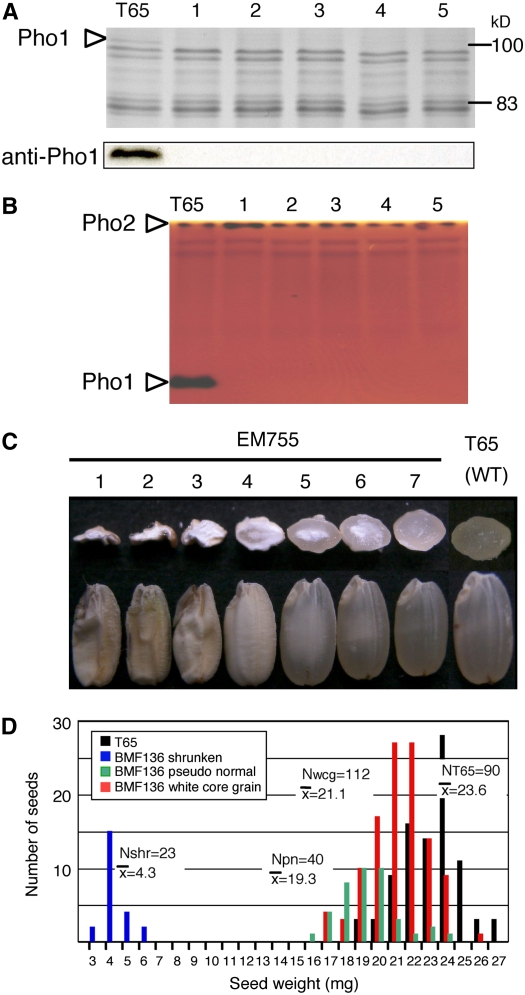
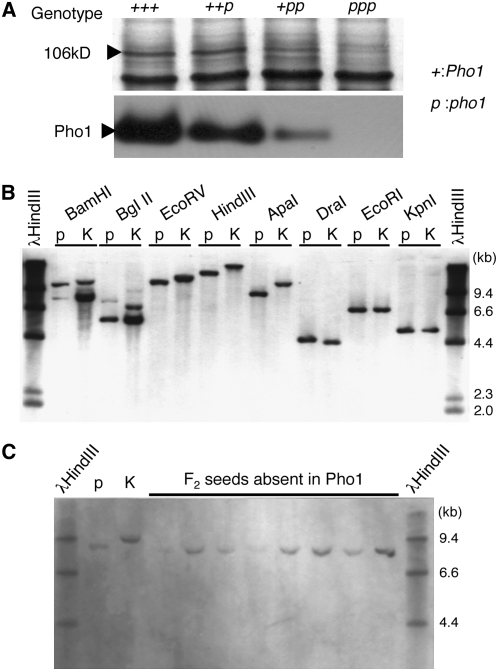
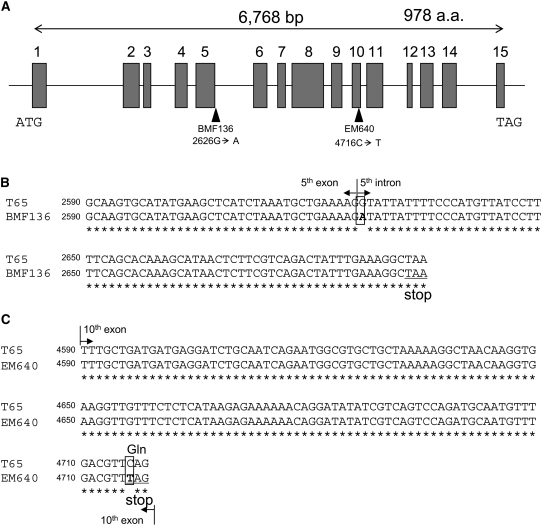
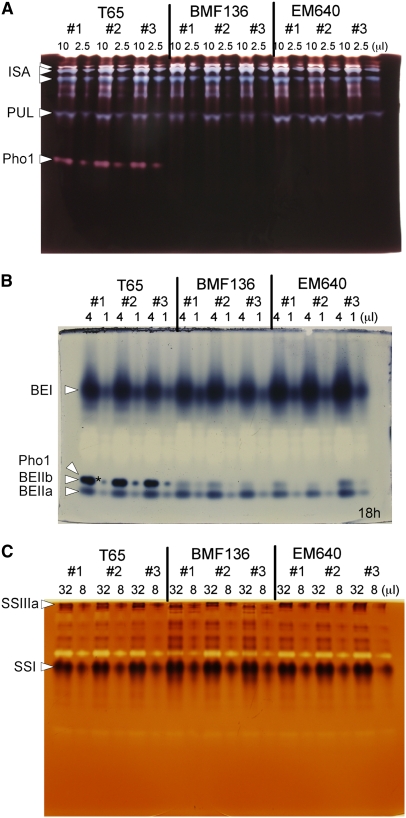
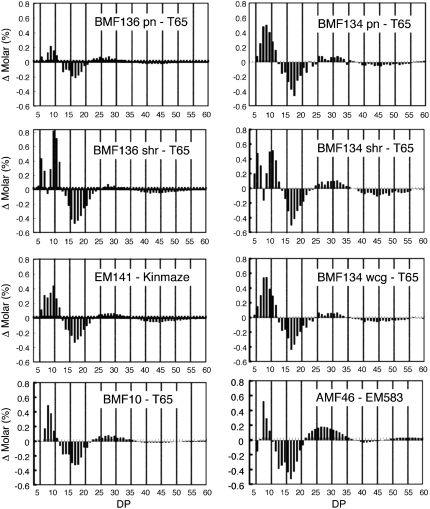
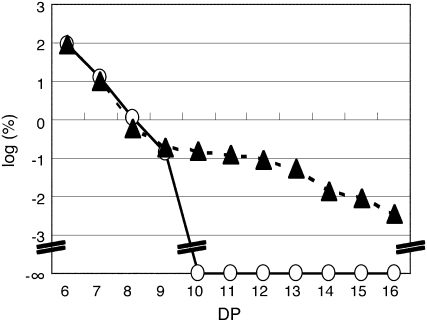
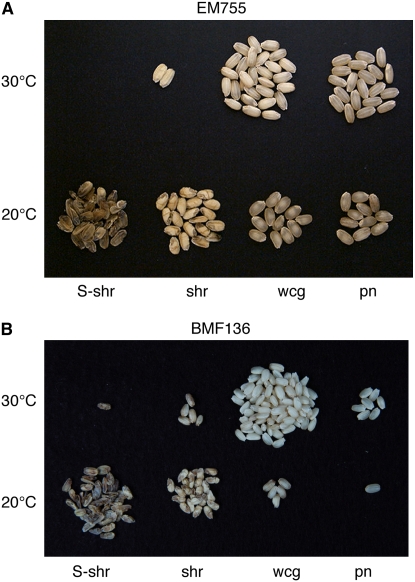
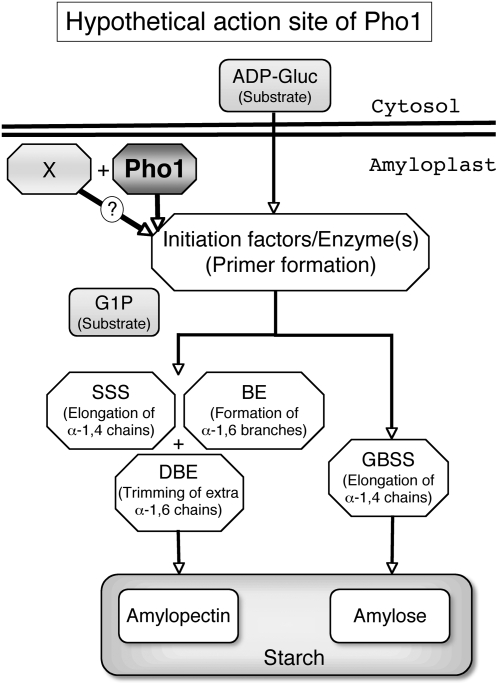
Plastidial phosphorylase (Pho1) accounts for approximately 96% of the total phosphorylase activity in developing rice (Oryza sativa) seeds. From mutant stocks induced by N-methyl-N-nitrosourea treatment, we identified plants with mutations in the Pho1 gene that are deficient in Pho1. Strikingly, the size of mature seeds and the starch content in these mutants showed considerable variation, ranging from shrunken to pseudonormal. The loss of Pho1 caused smaller starch granules to accumulate and modified the amylopectin structure. Variation in the morphological and biochemical phenotype of individual seeds was common to all 15 pho1-independent homozygous mutant lines studied, indicating that this phenotype was caused solely by the genetic defect. The phenotype of the pho1 mutation was temperature dependent. While the mutant plants grown at 30 degrees C produced mainly plump seeds at maturity, most of the seeds from plants grown at 20 degrees C were shrunken, with a significant proportion showing severe reduction in starch accumulation. These results strongly suggest that Pho1 plays a crucial role in starch biosynthesis in rice endosperm at low temperatures and that one or more other factors can complement the function of Pho1 at high temperatures.
Figures

References
-
- Ball, S.G., and Morell, M.K. (2003). From bacterial glycogen to starch: Understanding the biogenesis of the plant starch granule. Annu. Rev. Plant Biol. 54 207–233. - PubMed
-
- Borovsky, D., Smith, E.E., and Whelan, W.J. (1976). On the mechanism of amylose branching by potato Q-enzyme. Eur. J. Biochem. 62 307–312. - PubMed
-
- Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72 248–254. - PubMed
-
- Brown, J.W. (1996). Arabidopsis intron mutations and pre-mRNA splicing. Plant J. 10 771–780. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
