

Polytene chromosomal maps of 11 Drosophila species: the order of genomic scaffolds inferred from genetic and physical maps
- PMID: 18622037
- PMCID: PMC2475758
- DOI: 10.1534/genetics.107.086074
Polytene chromosomal maps of 11 Drosophila species: the order of genomic scaffolds inferred from genetic and physical maps
Abstract
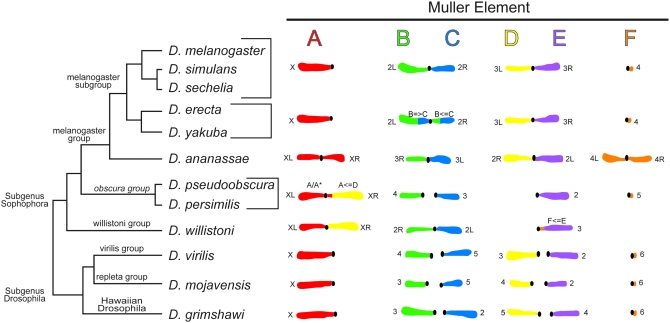
The sequencing of the 12 genomes of members of the genus Drosophila was taken as an opportunity to reevaluate the genetic and physical maps for 11 of the species, in part to aid in the mapping of assembled scaffolds. Here, we present an overview of the importance of cytogenetic maps to Drosophila biology and to the concepts of chromosomal evolution. Physical and genetic markers were used to anchor the genome assembly scaffolds to the polytene chromosomal maps for each species. In addition, a computational approach was used to anchor smaller scaffolds on the basis of the analysis of syntenic blocks. We present the chromosomal map data from each of the 11 sequenced non-Drosophila melanogaster species as a series of sections. Each section reviews the history of the polytene chromosome maps for each species, presents the new polytene chromosome maps, and anchors the genomic scaffolds to the cytological maps using genetic and physical markers. The mapping data agree with Muller's idea that the majority of Drosophila genes are syntenic. Despite the conservation of genes within homologous chromosome arms across species, the karyotypes of these species have changed through the fusion of chromosomal arms followed by subsequent rearrangement events.
Figures























Similar articles
-
Chromosomal rearrangement inferred from comparisons of 12 Drosophila genomes.Genetics. 2008 Jul;179(3):1657-80. doi: 10.1534/genetics.107.086108. Epub 2008 Jul 13. Genetics. 2008. PMID: 18622036 Free PMC article.
-
Cytological heterogeneity of heterochromatin among 10 sequenced Drosophila species.Genetics. 2022 Sep 30;222(2):iyac119. doi: 10.1093/genetics/iyac119. Genetics. 2022. PMID: 35946576 Free PMC article.
-
Reassignment of Drosophila willistoni Genome Scaffolds to Chromosome II Arms.G3 (Bethesda). 2015 Oct 4;5(12):2559-66. doi: 10.1534/g3.115.021311. G3 (Bethesda). 2015. PMID: 26438290 Free PMC article.
-
Mitotic and polytene chromosomes: comparisons between Drosophila melanogaster and Drosophila simulans.Genetica. 2004 Mar;120(1-3):137-50. doi: 10.1023/b:gene.0000017637.10230.c4. Genetica. 2004. PMID: 15088654 Review.
-
Mapping Drosophila genomic aberration breakpoints with comparative genome hybridization on microarrays.Methods Enzymol. 2006;410:377-86. doi: 10.1016/S0076-6879(06)10018-X. Methods Enzymol. 2006. PMID: 16938561 Review.
Cited by
-
Comparative Genomic Analyses Provide New Insights into the Evolutionary Dynamics of Heterochromatin in Drosophila.PLoS Genet. 2016 Aug 11;12(8):e1006212. doi: 10.1371/journal.pgen.1006212. eCollection 2016 Aug. PLoS Genet. 2016. PMID: 27513559 Free PMC article.
-
FISH mapping of microsatellite loci from Drosophila subobscura and its comparison to related species.Chromosome Res. 2010 Feb;18(2):213-26. doi: 10.1007/s10577-010-9112-4. Epub 2010 Mar 3. Chromosome Res. 2010. PMID: 20198419
-
Assessing the Architecture of Drosophila mojavensis Locomotor Evolution with Bulk Segregant Analysis.G3 (Bethesda). 2019 May 7;9(5):1767-1775. doi: 10.1534/g3.119.400036. G3 (Bethesda). 2019. PMID: 30926724 Free PMC article.
-
Divergence of X-linked trans regulatory proteins and the misexpression of gene targets in sterile Drosophila pseudoobscura hybrids.BMC Genomics. 2022 Jan 6;23(1):30. doi: 10.1186/s12864-021-08267-w. BMC Genomics. 2022. PMID: 34991488 Free PMC article.
-
Chromosomes as Barcodes: Discovery of a New Species of Black Fly (Diptera: Simuliidae) from California, USA.Insects. 2022 Oct 4;13(10):903. doi: 10.3390/insects13100903. Insects. 2022. PMID: 36292851 Free PMC article.
References
-
- Anderson, W. W., F. J. Ayala and R. E. Michod, 1977. Chromosomal and allozymic diagnosis of three species of Drosophila, Drosophila pseudoobscura, Drosophila persimilis, and Drosophila miranda. J. Hered. 68 71–74. - PubMed
-
- Ashburner, M., and F. Lemeunier, 1976. Relationships within the melanogaster species subgroup of the genus Drosophila (Sophophora). I. Inversion polymorphisms in Drosophila melanogaster and Drosophila simulans. Proc. R. Soc. Lond. B Biol. Sci. 193 137–157. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
