

Immune-driven recombination and loss of control after HIV superinfection
- PMID: 18625749
- PMCID: PMC2525594
- DOI: 10.1084/jem.20080281
Immune-driven recombination and loss of control after HIV superinfection
Abstract
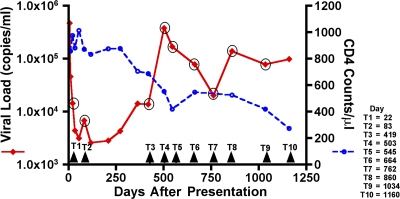
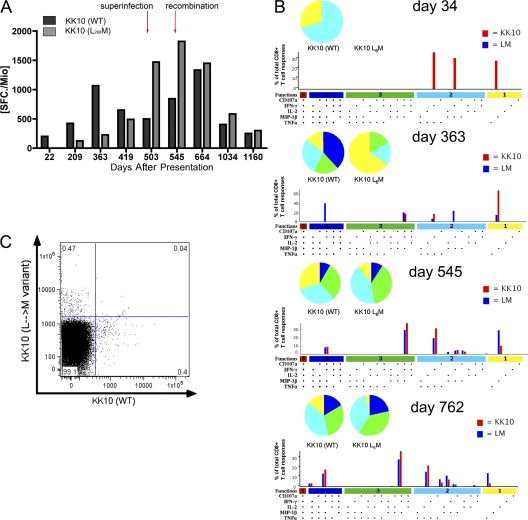
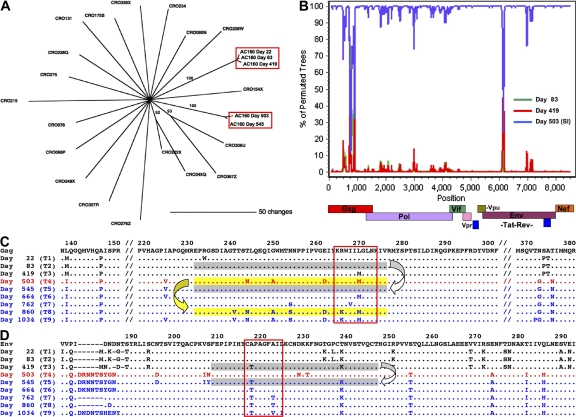
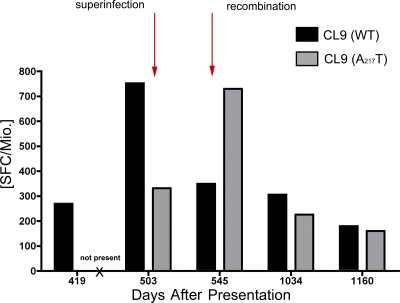
After acute HIV infection, CD8(+) T cells are able to control viral replication to a set point. This control is often lost after superinfection, although the mechanism behind this remains unclear. In this study, we illustrate in an HLA-B27(+) subject that loss of viral control after HIV superinfection coincides with rapid recombination events within two narrow regions of Gag and Env. Screening for CD8(+) T cell responses revealed that each of these recombination sites (approximately 50 aa) encompassed distinct regions containing two immunodominant CD8 epitopes (B27-KK10 in Gag and Cw1-CL9 in Env). Viral escape and the subsequent development of variant-specific de novo CD8(+) T cell responses against both epitopes were illustrative of the significant immune selection pressures exerted by both responses. Comprehensive analysis of the kinetics of CD8 responses and viral evolution indicated that the recombination events quickly facilitated viral escape from both dominant WT- and variant-specific responses. These data suggest that the ability of a superinfecting strain of HIV to overcome preexisting immune control may be related to its ability to rapidly recombine in critical regions under immune selection pressure. These data also support a role for cellular immune pressures in driving the selection of new recombinant forms of HIV.
Figures
References
-
- Schmitz, J.E., M.J. Kuroda, S. Santra, V.G. Sasseville, M.A. Simon, M.A. Lifton, P. Racz, K. Tenner-Racz, M. Dalesandro, B.J. Scallon, et al. 1999. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science. 283:857–860. - PubMed
-
- Carrington, M., and S.J. O'Brien. 2003. The influence of HLA genotype on AIDS. Annu. Rev. Med. 54:535–551. - PubMed
-
- Streeck, H., M. Lichterfeld, G. Alter, A. Meier, N. Teigen, B. Yassine-Diab, H.K. Sidhu, S. Little, A. Kelleher, J.P. Routy, et al. 2007. Recognition of a defined region within p24 gag by CD8+ T cells during primary human immunodeficiency virus type 1 infection in individuals expressing protective HLA class I alleles. J. Virol. 81:7725–7731. - PMC - PubMed
-
- Leslie, A.J., K.J. Pfafferott, P. Chetty, R. Draenert, M.M. Addo, M. Feeney, Y. Tang, E.C. Holmes, T. Allen, J.G. Prado, et al. 2004. HIV evolution: CTL escape mutation and reversion after transmission. Nat. Med. 10:282–289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
