

Mars promotes dTACC dephosphorylation on mitotic spindles to ensure spindle stability
- PMID: 18625841
- PMCID: PMC2447907
- DOI: 10.1083/jcb.200712080
Mars promotes dTACC dephosphorylation on mitotic spindles to ensure spindle stability
Abstract
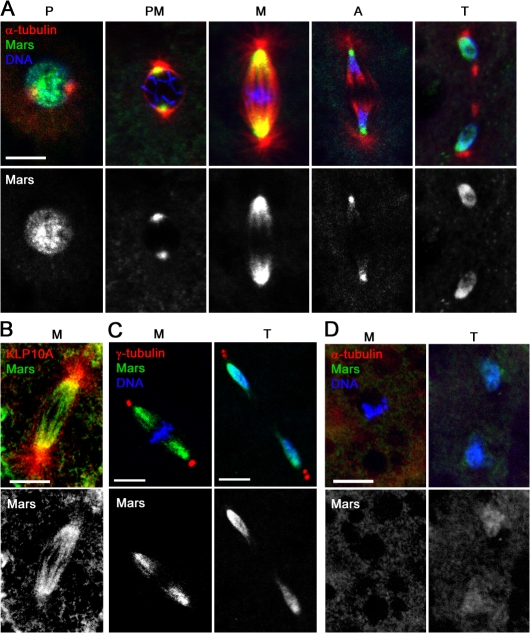
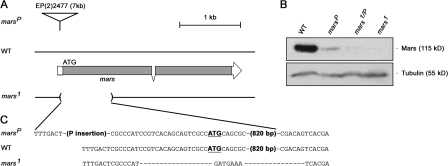
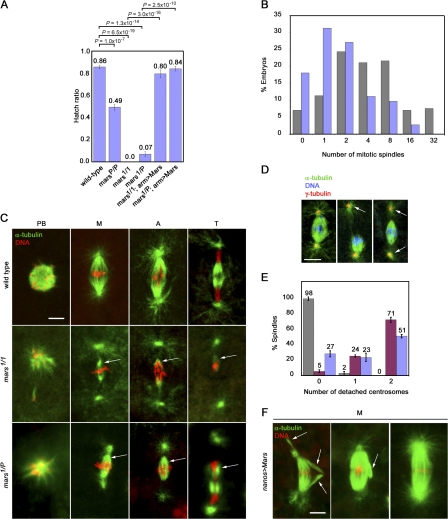
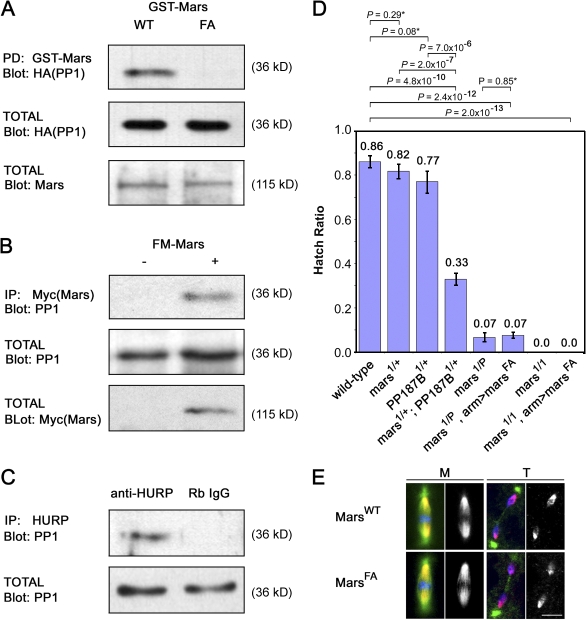
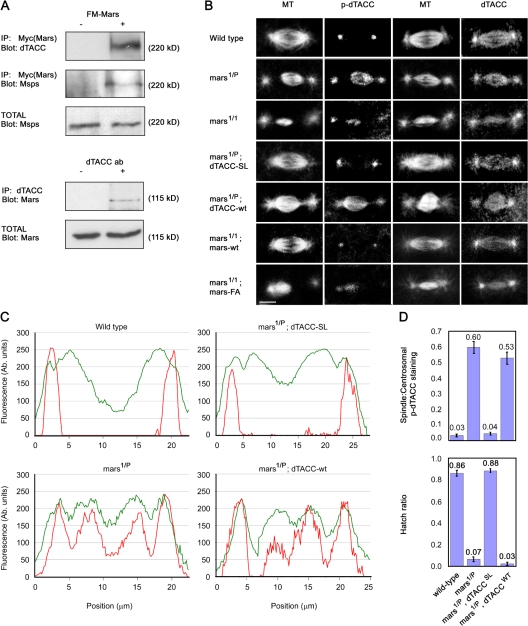
Microtubule-associated proteins (MAPs) ensure the fidelity of chromosome segregation by controlling microtubule (MT) dynamics and mitotic spindle stability. However, many aspects of MAP function and regulation are poorly understood in a developmental context. We show that mars, which encodes a Drosophila melanogaster member of the hepatoma up-regulated protein family of MAPs, is essential for MT stabilization during early embryogenesis. As well as associating with spindle MTs in vivo, Mars binds directly to protein phosphatase 1 (PP1) and coimmunoprecipitates from embryo extracts with minispindles and Drosophila transforming acidic coiled-coil (dTACC), two MAPs that function as spindle assembly factors. Disruption of binding to PP1 or loss of mars function results in elevated levels of phosphorylated dTACC on spindles. A nonphosphorylatable form of dTACC is capable of rescuing the lethality of mars mutants. We propose that Mars mediates spatially controlled dephosphorylation of dTACC, which is critical for spindle stabilization.
Figures
References
-
- Baksa, K., H. Morawietz, V. Dombrádi, M. Axton, H. Taubert, G. Szabo, I. Torok, A. Udvardy, H. Gyurkovics, and B. Szöor. 1993. Mutations in the protein phosphatase 1 gene at 87B can differentially affect suppression of position-effect variegation and mitosis in Drosophila melanogaster.Genetics. 135:117–125. - PMC - PubMed
-
- Bennett, D., and L. Alphey. 2004. Cloning and expression of mars, a novel member of the guanylate kinase associated protein family in Drosophila. Gene Expr. Patterns. 4:529–535. - PubMed
-
- Bennett, D., E. Lyulcheva, and L. Alphey. 2006. Towards a comprehensive analysis of the protein phosphatase 1 interactome in Drosophila. J. Mol. Biol. 364:196–212. - PubMed
-
- Brittle, A.L., and H. Ohkura. 2005. Centrosome maturation: Aurora lights the way to the poles. Curr. Biol. 15:R880–R882. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
