

Palatal seam disintegration: to die or not to die? that is no longer the question
- PMID: 18629865
- PMCID: PMC2659566
- DOI: 10.1002/dvdy.21599
Palatal seam disintegration: to die or not to die? that is no longer the question
Abstract
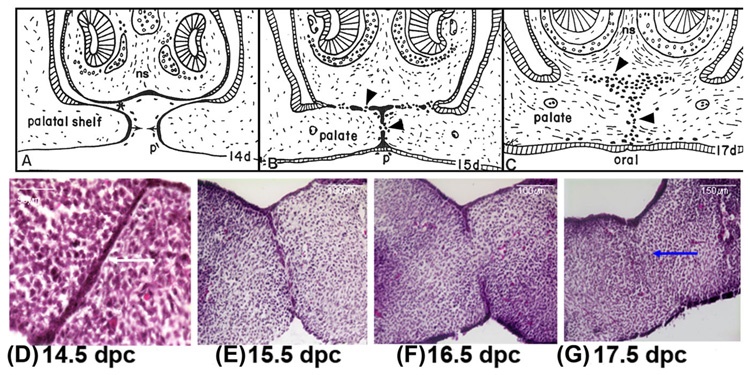
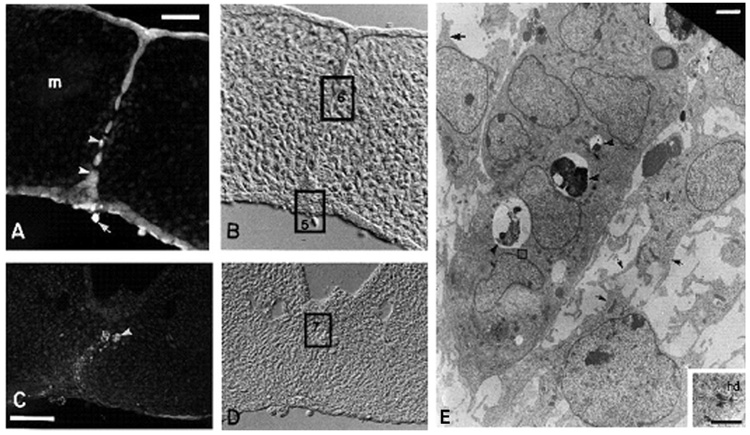
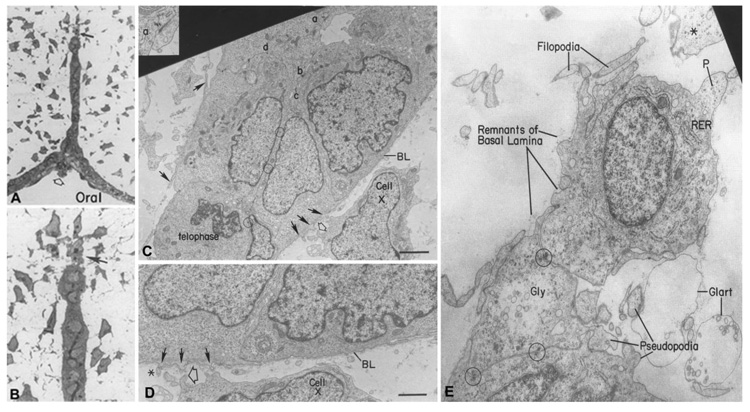
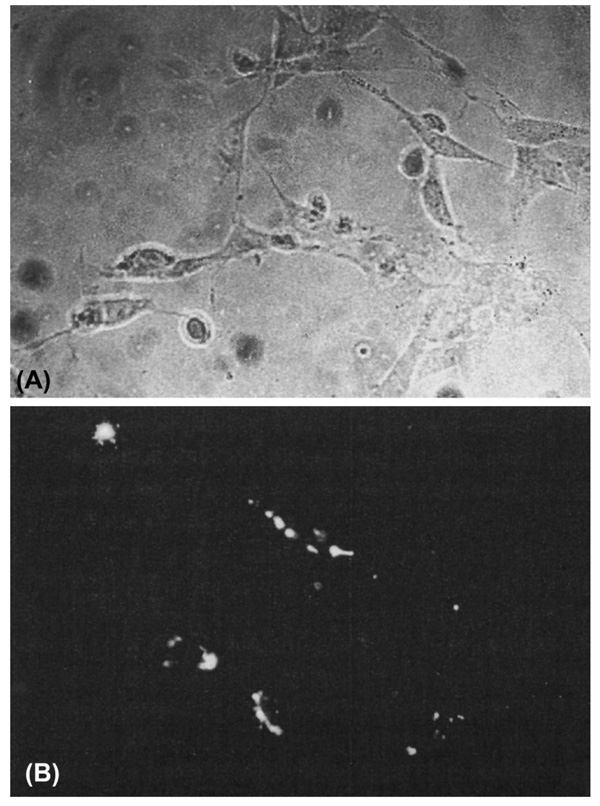
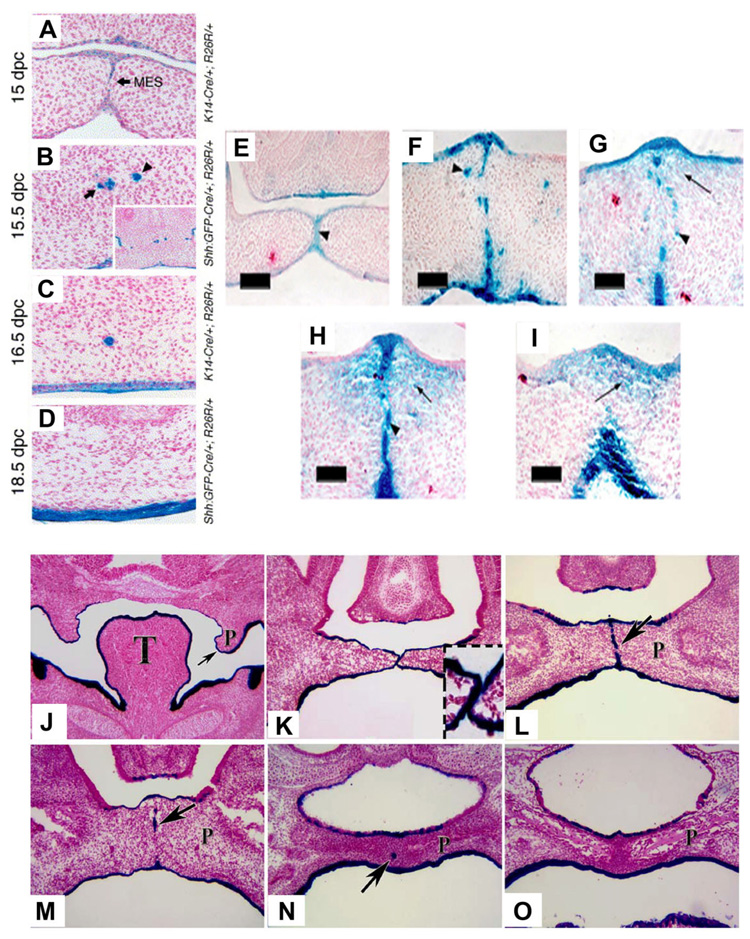
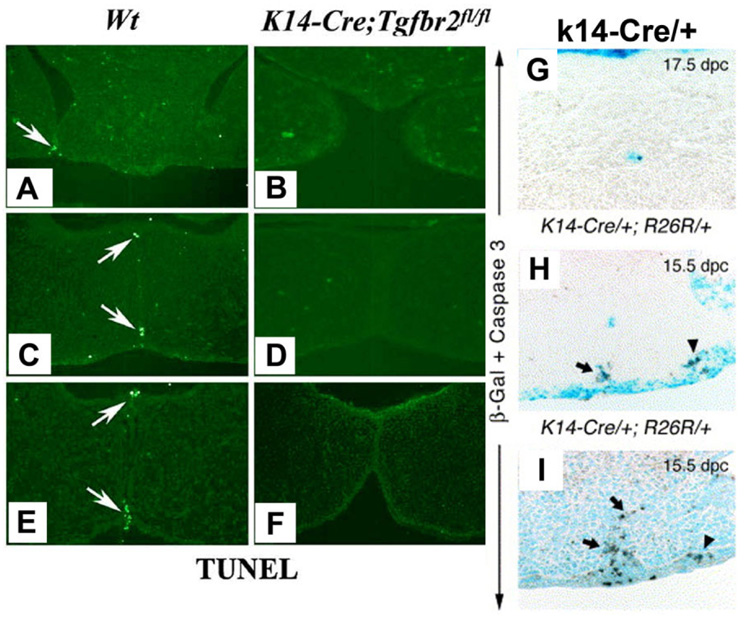
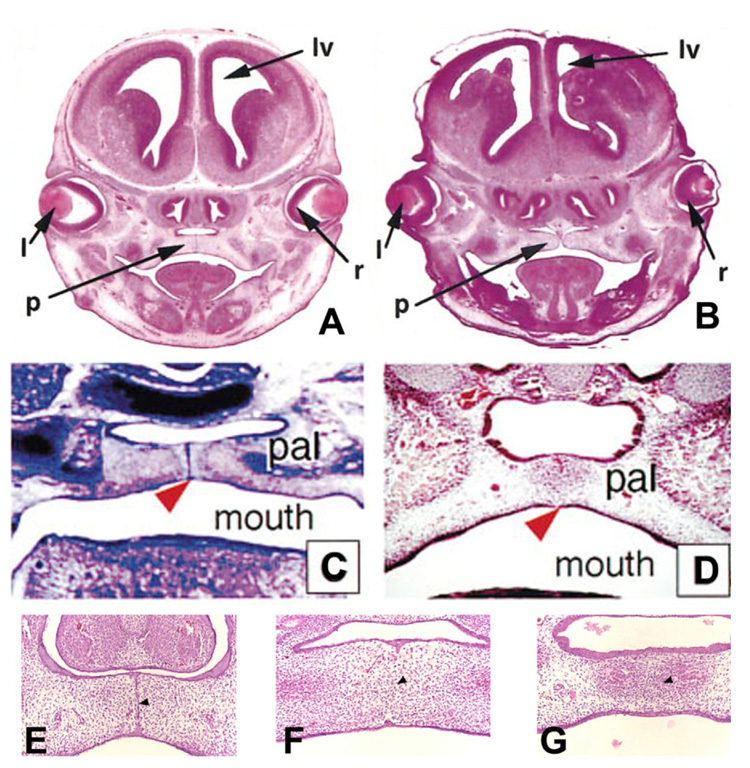
Formation of the medial epithelial seam (MES) by palatal shelf fusion is a crucial step of palate development. Complete disintegration of the MES is the final essential phase of palatal confluency with surrounding mesenchymal cells. In general, the mechanisms of palatal seam disintegration are not overwhelmingly complex, but given the large number of interacting constituents; their complicated circuitry involving feedforward, feedback, and crosstalk; and the fact that the kinetics of interaction matter, this otherwise simple mechanism can be quite difficult to interpret. As a result of this complexity, apparently simple but highly important questions remain unanswered. One such question pertains to the fate of the palatal seam. Such questions may be answered by detailed and extensive quantitative experimentation of basic biological studies (cellular, structural) and the newest molecular biological determinants (genetic/dye cell lineage, gene activity, kinase/enzyme activity), as well as animal model (knockouts, transgenic) approaches. System biology and cellular kinetics play a crucial role in cellular MES function; omissions of such critical contributors may lead to inaccurate understanding of the fate of MES. Excellent progress has been made relevant to elucidation of the mechanism(s) of palatal seam disintegration. Current understanding of palatal seam disintegration suggests epithelial-mesenchymal transition and/or programmed cell death as two most common mechanisms of MES disintegration. In this review, I discuss those two mechanisms and the differences between them.
Copyright (c) 2008 Wiley-Liss, Inc.
Figures
References
-
- Abdelilah-Seyfried S, Cox DN, Jan YN. Bazooka is a permissive factor for the invasive behavior of discs large tumor cells in Drosophila ovarian follicular epithelia. Development. 2003;130:1927–1935. - PubMed
-
- Abraham MC, Shaham S. Death without caspases, caspases without death. Trends Cell Biol. 2004;14:184–193. - PubMed
-
- Barasch J. Genes and proteins involved in mesenchymal to epithelial transition. Curr Opin Nephrol Hypertens. 2001;10:429–436. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
