

Reprogramming multipotent tumor cells with the embryonic neural crest microenvironment
- PMID: 18629870
- PMCID: PMC2570047
- DOI: 10.1002/dvdy.21613
Reprogramming multipotent tumor cells with the embryonic neural crest microenvironment
Abstract
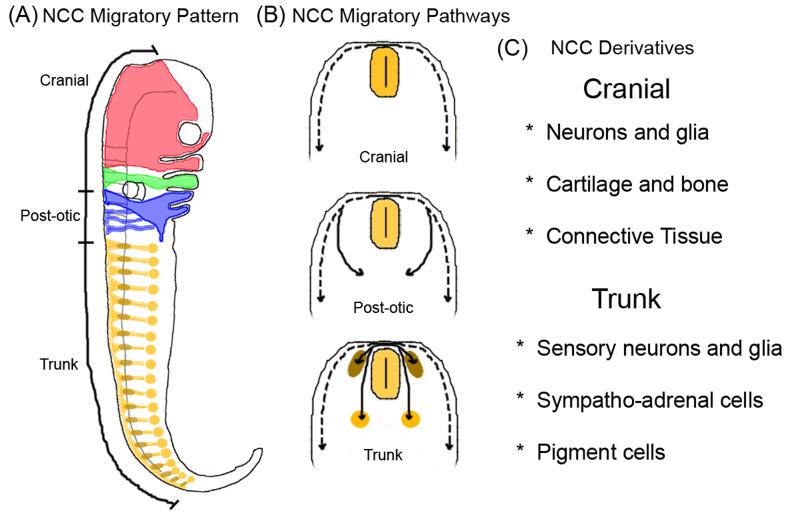
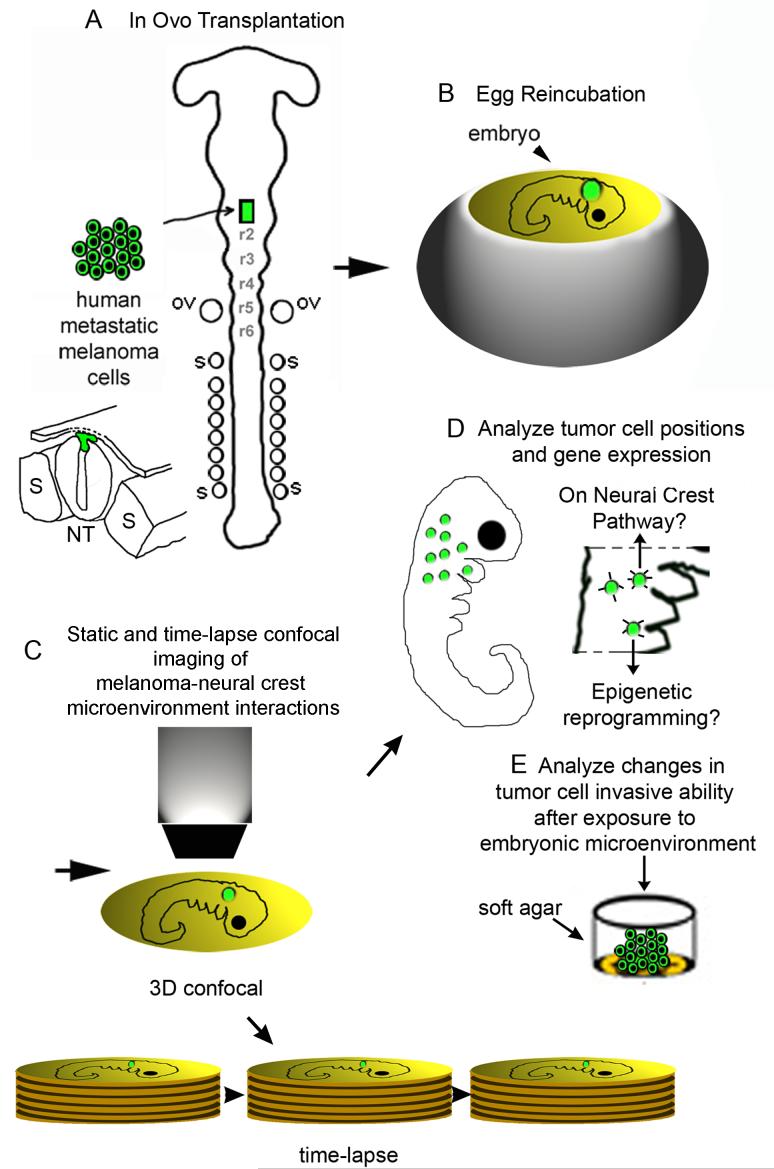
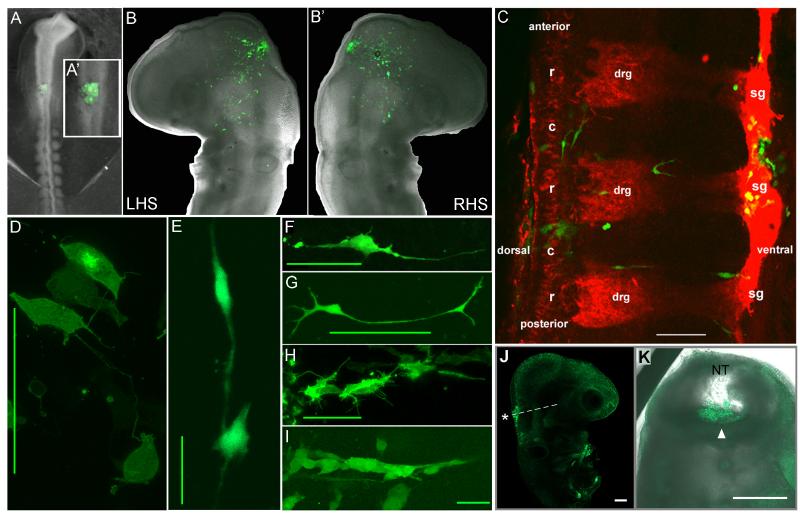
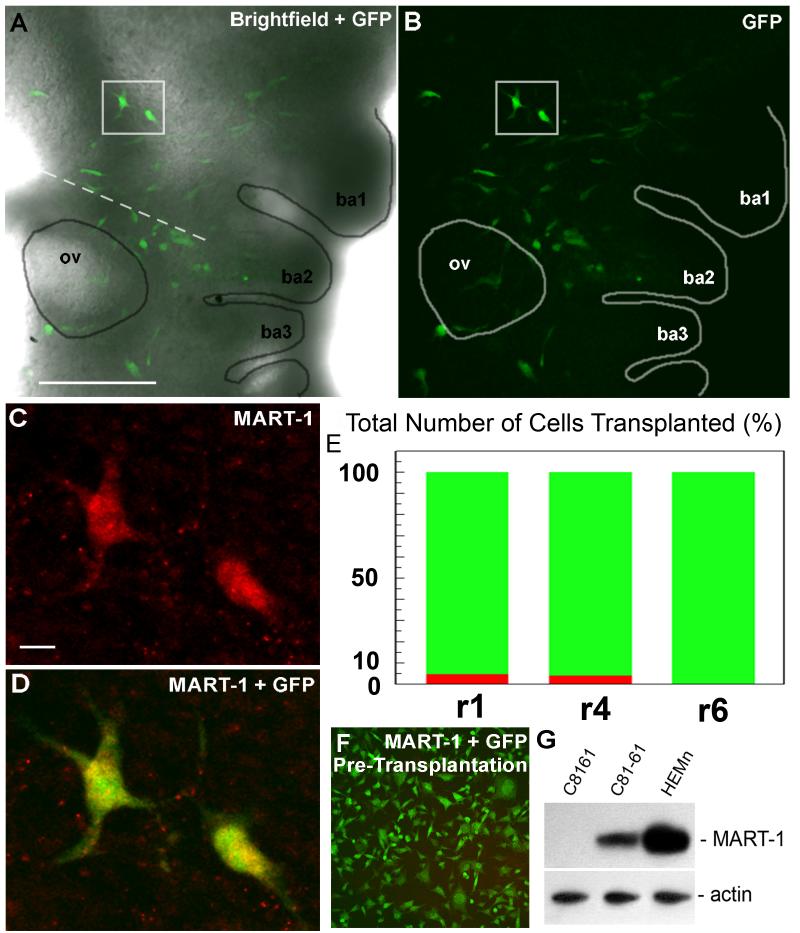
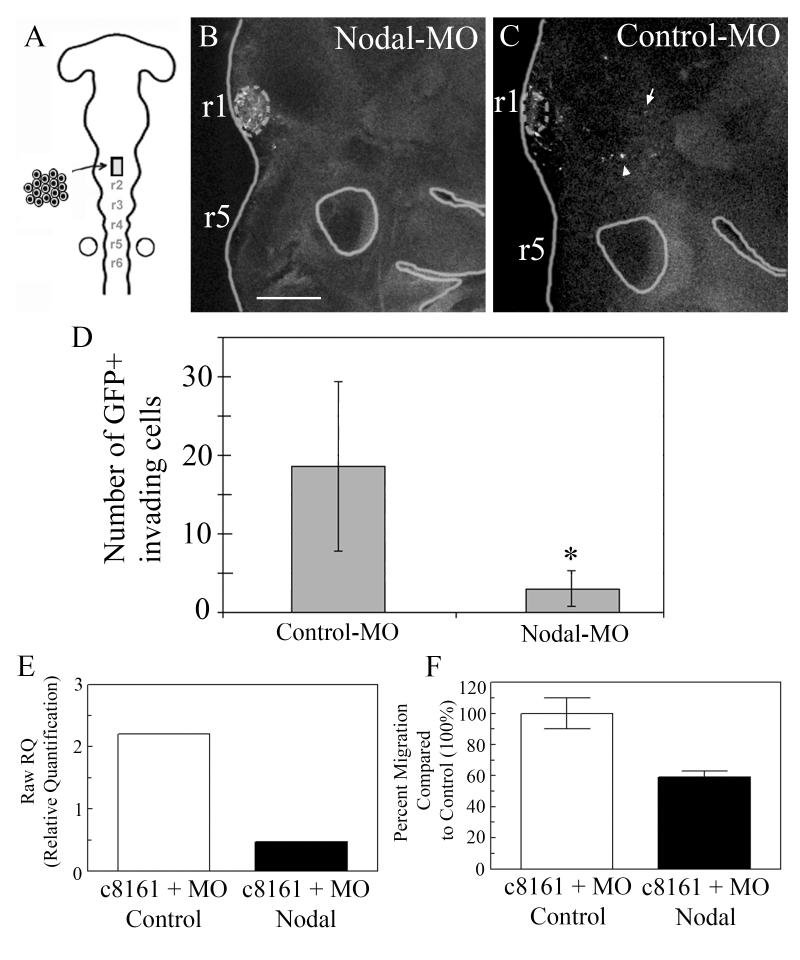
The embryonic microenvironment is an important source of signals that program multipotent cells to adopt a particular fate and migratory path, yet its potential to reprogram and restrict multipotent tumor cell fate and invasion is unrealized. Aggressive tumor cells share many characteristics with multipotent, invasive embryonic progenitors, contributing to the paradigm of tumor cell plasticity. In the vertebrate embryo, multiple cell types originate from a highly invasive cell population called the neural crest. The neural crest and the embryonic microenvironments they migrate through represent an excellent model system to study cell diversification during embryogenesis and phenotype determination. Recent exciting studies of tumor cells transplanted into various embryo models, including the neural crest rich chick microenvironment, have revealed the potential to control and revert the metastatic phenotype, suggesting further work may help to identify new targets for therapeutic intervention derived from a convergence of tumorigenic and embryonic signals. In this mini-review, we summarize markers that are common to the neural crest and highly aggressive human melanoma cells. We highlight advances in our understanding of tumor cell behaviors and plasticity studied within the chick neural crest rich microenvironment. In so doing, we honor the tremendous contributions of Professor Elizabeth D. Hay toward this important interface of developmental and cancer biology.
Copyright (c) 2008 Wiley-Liss, Inc.
Figures
References
-
- Anderson RB, Newgreen DF, Young HM. Neural crest and the development of the enteric nervous system. Adv. Exp. Med. Biol. 2006;589:181–96. - PubMed
-
- Barrallo-Gimeno A, Nieto MA. The Snail genes as inducers of cell movement and survival: implications in development and cancer. Dev. 2005;132:3151–3161. - PubMed
-
- Bilozur ME, Hay ED. Neural crest migration in 3D extracellular matrix utilizes laminin, fibronectin, or collagen. Dev Biol. 1988;125:19–33. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
