

Functional topography and integration of the contralateral and ipsilateral retinocollicular projections of ephrin-A-/- mice
- PMID: 18632942
- PMCID: PMC6670399
- DOI: 10.1523/JNEUROSCI.1135-08.2008
Functional topography and integration of the contralateral and ipsilateral retinocollicular projections of ephrin-A-/- mice
Abstract
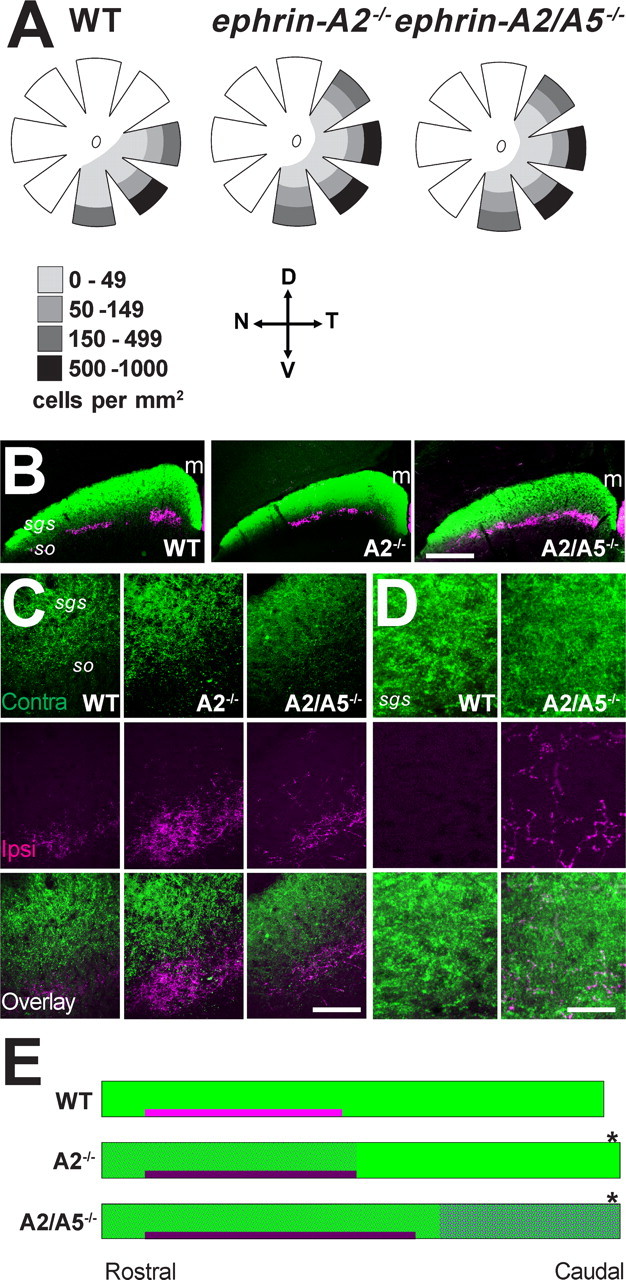
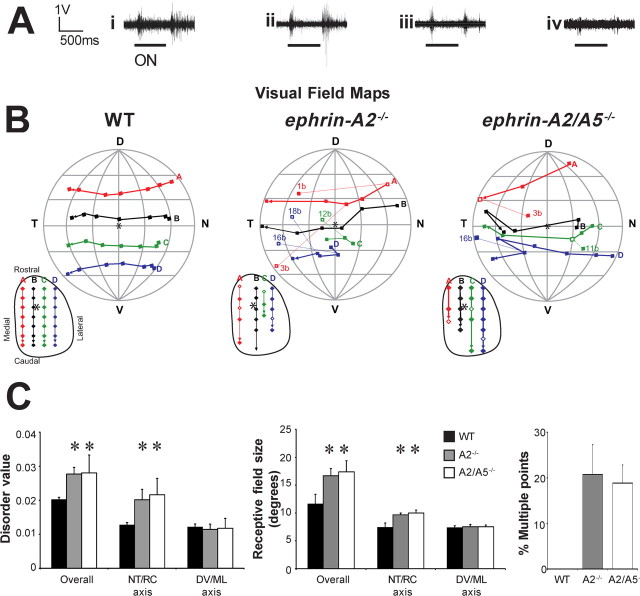
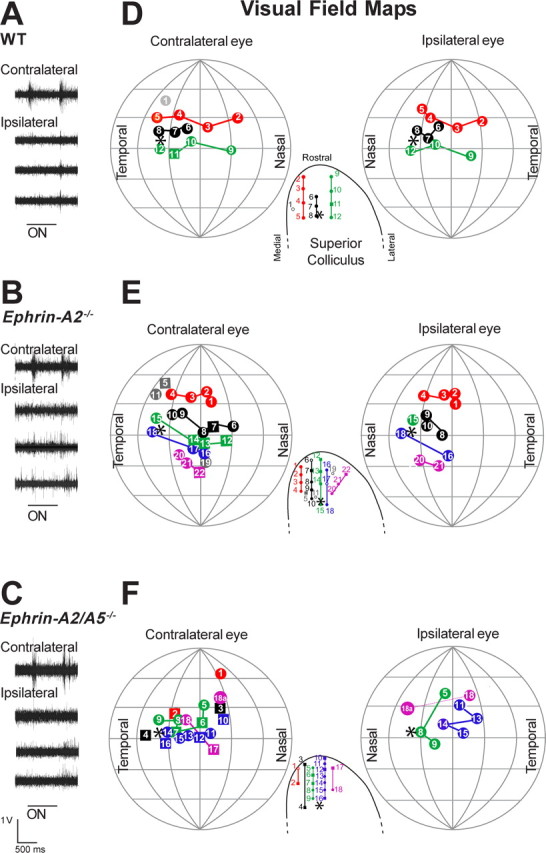
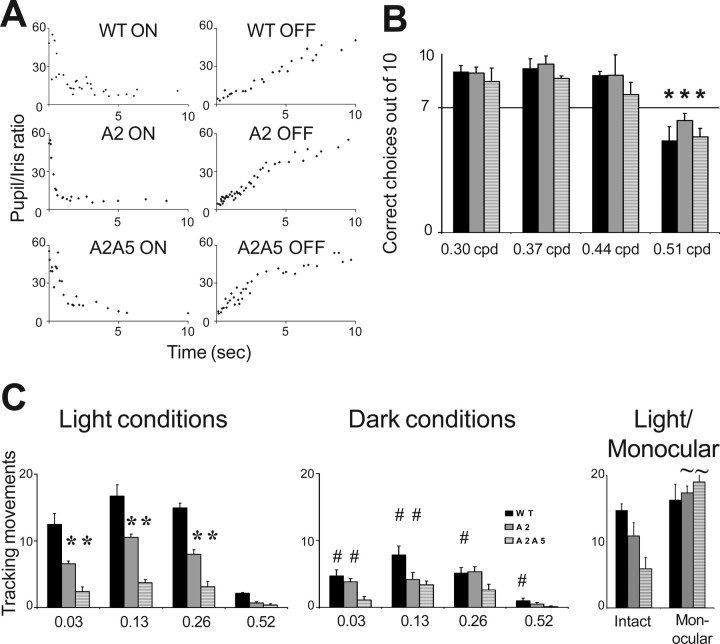
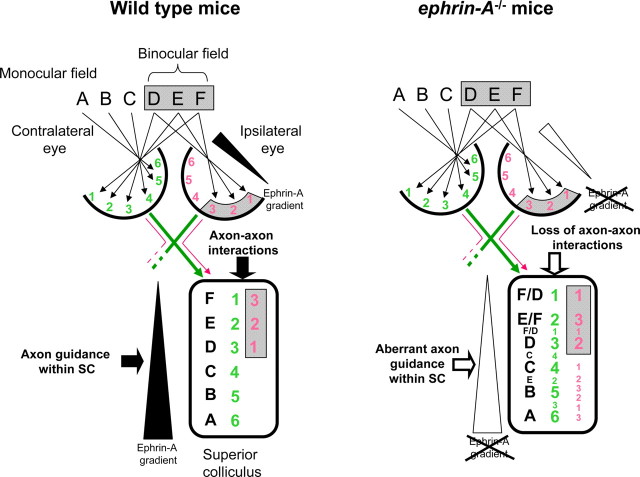
Topographically ordered projections are established by molecular guidance cues and refined by neuronal activity. Retinal input to a primary visual center, the superior colliculus (SC), is bilateral with a dense contralateral projection and a sparse ipsilateral one. Both projections are topographically organized, but in opposing anterior-posterior orientations. This arrangement provides functionally coherent input to each colliculus from the binocular visual field, supporting visual function. When guidance cues involved in contralateral topography (ephrin-As) are absent, crossed retinal ganglion cell (RGC) axons form inappropriate terminations within the SC. However, the organization of the ipsilateral projection relative to the abnormal contralateral input remains unknown, as does the functional capacity of both projections. We show here that in ephrin-A(-/-) mice, the SC contains an expanded, diffuse ipsilateral projection. Electrophysiological recording demonstrated that topography of visually evoked responses recorded from the contralateral superior colliculus of ephrin-A(-/-) mice displayed similar functional disorder in all genotypes, contrasting with their different degrees of anatomical disorder. In contrast, ipsilateral responses were retinotopic in ephrin-A2(-/-) but disorganized in ephrin-A2/A5(-/-) mice. The lack of integration of binocular input resulted in specific visual deficits, which could be reversed by occlusion of one eye. The discrepancy between anatomical and functional topography in both the ipsilateral and contralateral projections implies suppression of inappropriately located terminals. Moreover, the misalignment of ipsilateral and contralateral visual information in ephrin-A2/A5(-/-) mice suggests a role for ephrin-As in integrating convergent visual inputs.
Figures
References
-
- Caras IW. A link between axon guidance and axon fasciculation suggested by studies of the tyrosine kinase receptor EphA5/REK7 and its ligand ephrin-A5/AL-1. Cell Tissue Res. 1997;290:261–264. - PubMed
-
- Cheng HJ, Nakamoto M, Bergemann AD, Flanagan JG. Complementary gradients in expression and binding of ELF-1 and Mek4 in development of the topographic retinotectal projection map. Cell. 1995;82:371–381. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials