

Bluetongue virus: dissection of the polymerase complex
- PMID: 18632949
- PMCID: PMC2735681
- DOI: 10.1099/vir.0.2008/002089-0
Bluetongue virus: dissection of the polymerase complex
Abstract
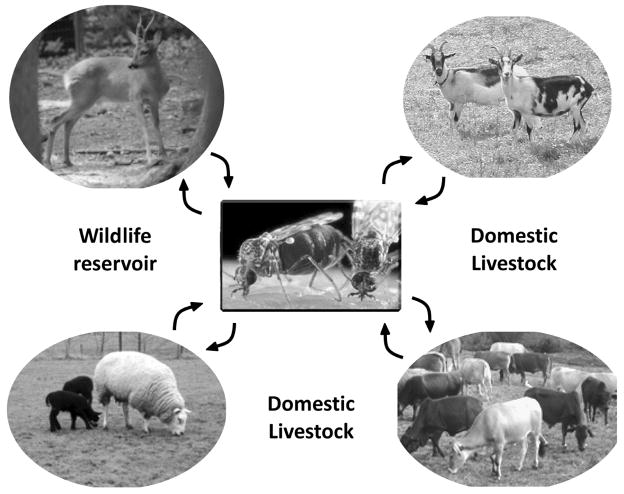
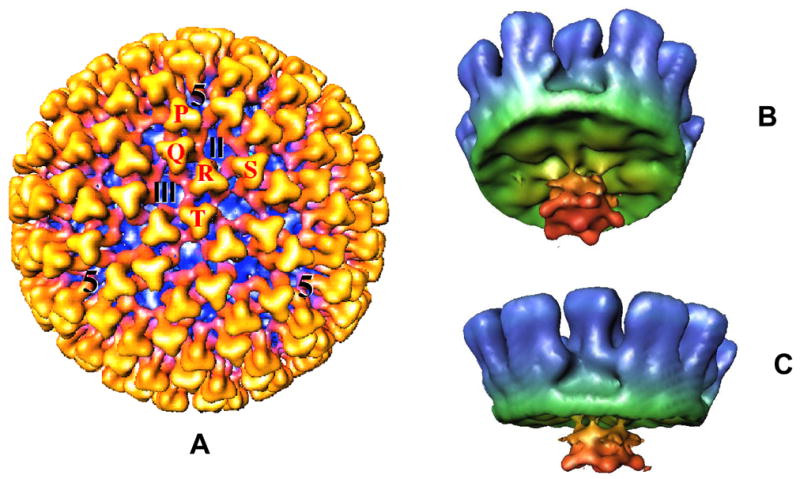
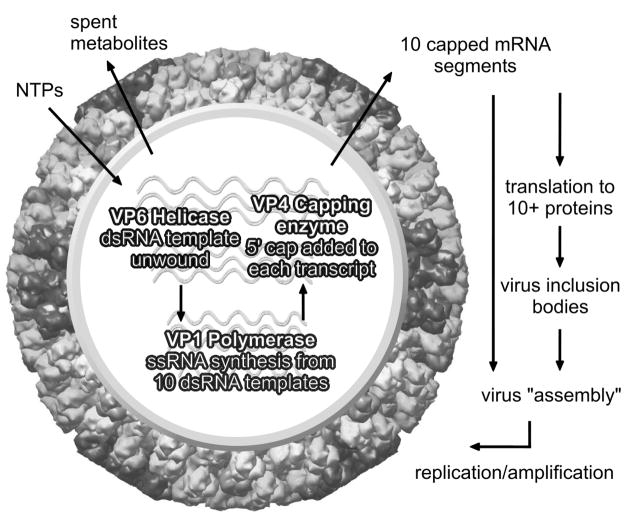
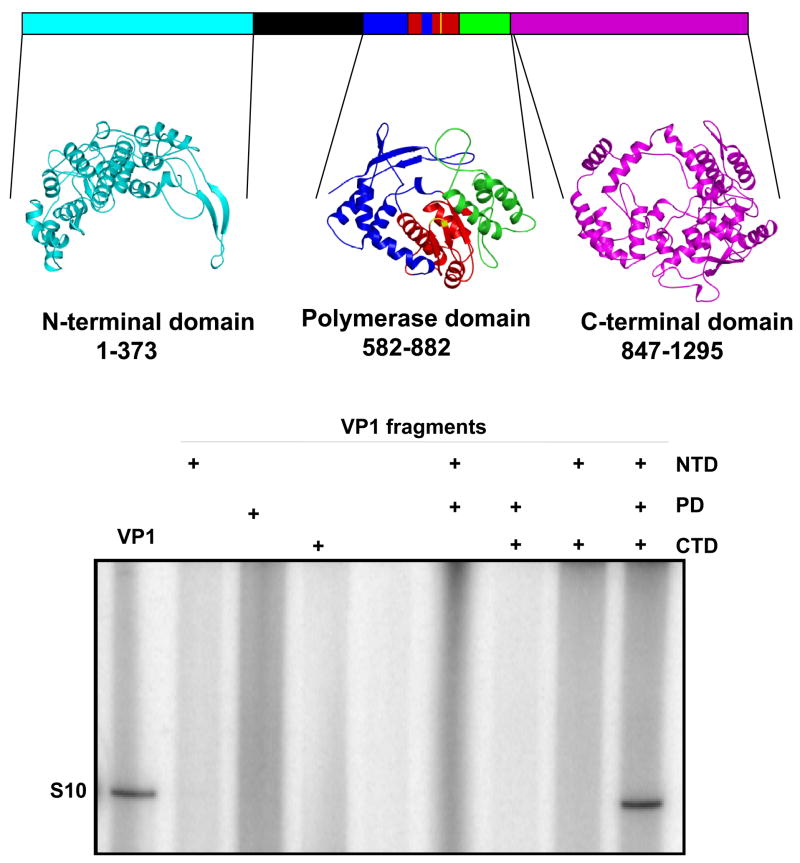
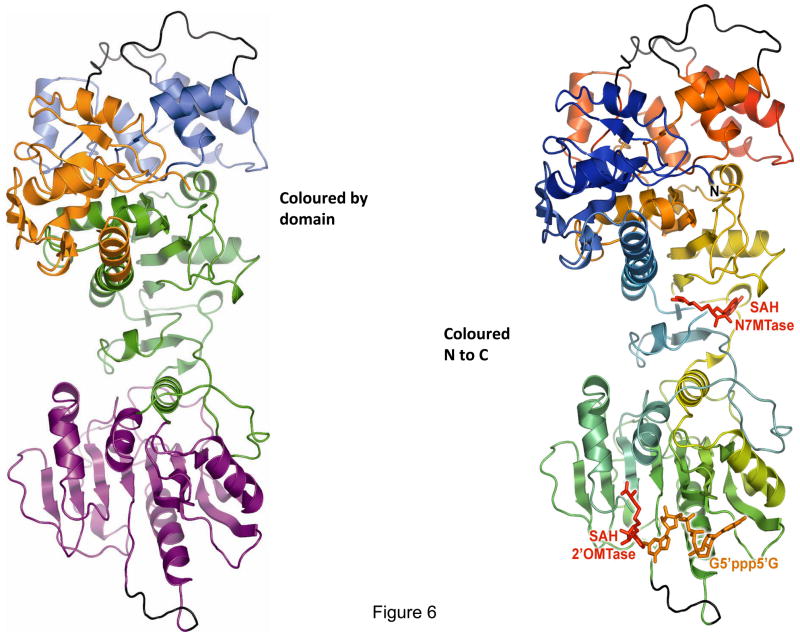
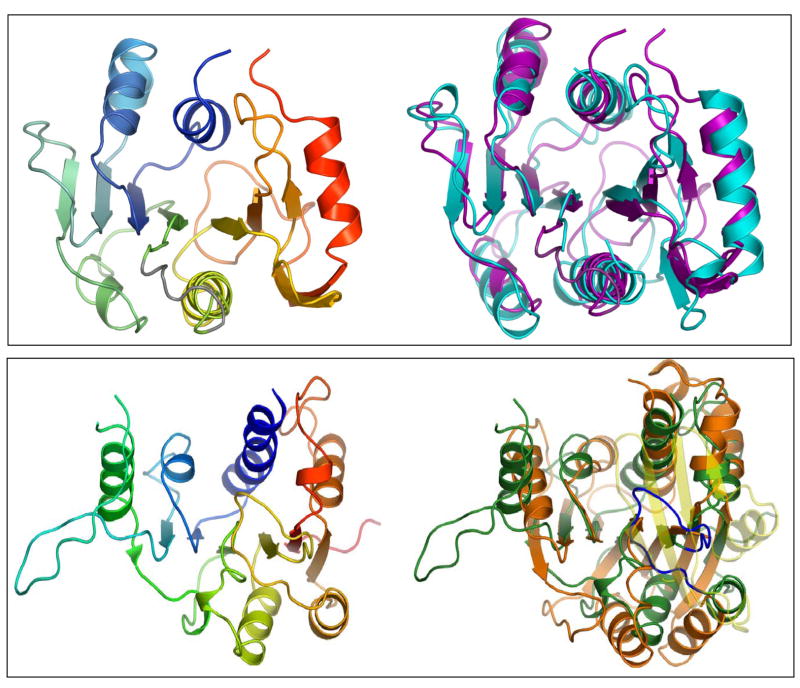
Bluetongue is a vector-borne viral disease of ruminants that is endemic in tropical and subtropical countries. Since 1998 the virus has also appeared in Europe. Partly due to the seriousness of the disease, bluetongue virus (BTV), a member of genus Orbivirus within the family Reoviridae, has been a subject of intense molecular study for the last three decades and is now one of the best understood viruses at the molecular and structural levels. BTV is a complex non-enveloped virus with seven structural proteins arranged in two capsids and a genome of ten double-stranded (ds) RNA segments. Shortly after cell entry, the outer capsid is lost to release an inner capsid (the core) which synthesizes capped mRNAs from each genomic segment, extruding them into the cytoplasm. This requires the efficient co-ordination of a number of enzymes, including helicase, polymerase and RNA capping activities. This review will focus on our current understanding of these catalytic proteins as derived from the use of recombinant proteins, combined with functional assays and the in vitro reconstitution of the transcription/replication complex. In some cases, 3D structures have complemented this analysis to reveal the fine structural detail of these proteins. The combined activities of the core enzymes produce infectious transcripts necessary and sufficient to initiate BTV infection. Such infectious transcripts can now be synthesized wholly in vitro and, when introduced into cells by transfection, lead to the recovery of infectious virus. Future studies thus hold the possibility of analysing the consequence of mutation in a replicating virus system.
Figures
References
-
- Ahnert P, Patel SS. Asymmetric interactions of hexameric bacteriophage T7 DNA helicase with the 5′- and 3′-tails of the forked DNA substrate. J Biol Chem. 1997;272:32267–32273. - PubMed
-
- Barbosa PR, Sousa MO, Barbosa EC, Bomfim Ade S, Ginefra P, Nadal J. Analysis of the prevalence of ventricular late potentials in the late phase of myocardial infarction based on the site of infarction. Arq Bras Cardiol. 2002;78:352–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources