

Unfolding and melting of DNA (RNA) hairpins: the concept of structure-specific 2D dynamic landscapes
- PMID: 18633543
- PMCID: PMC2548322
- DOI: 10.1039/b804675c
Unfolding and melting of DNA (RNA) hairpins: the concept of structure-specific 2D dynamic landscapes
Abstract
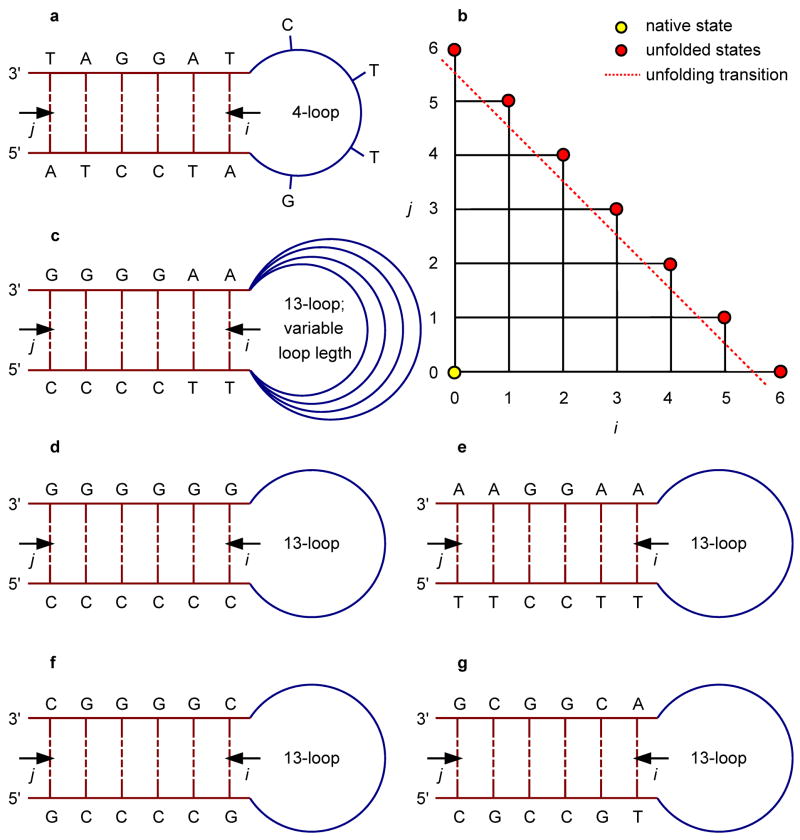
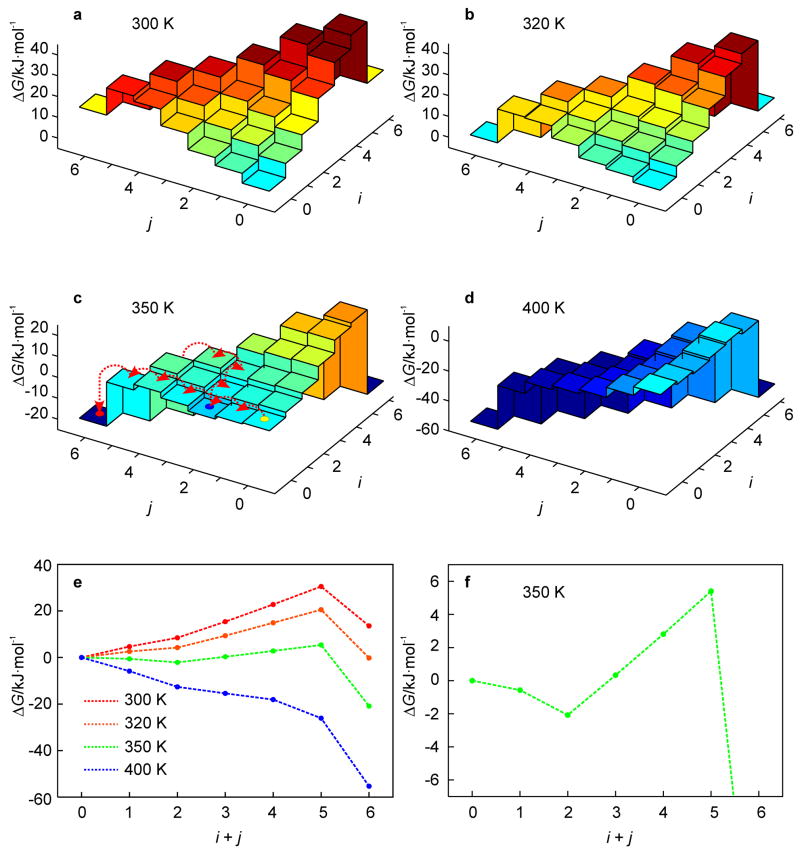
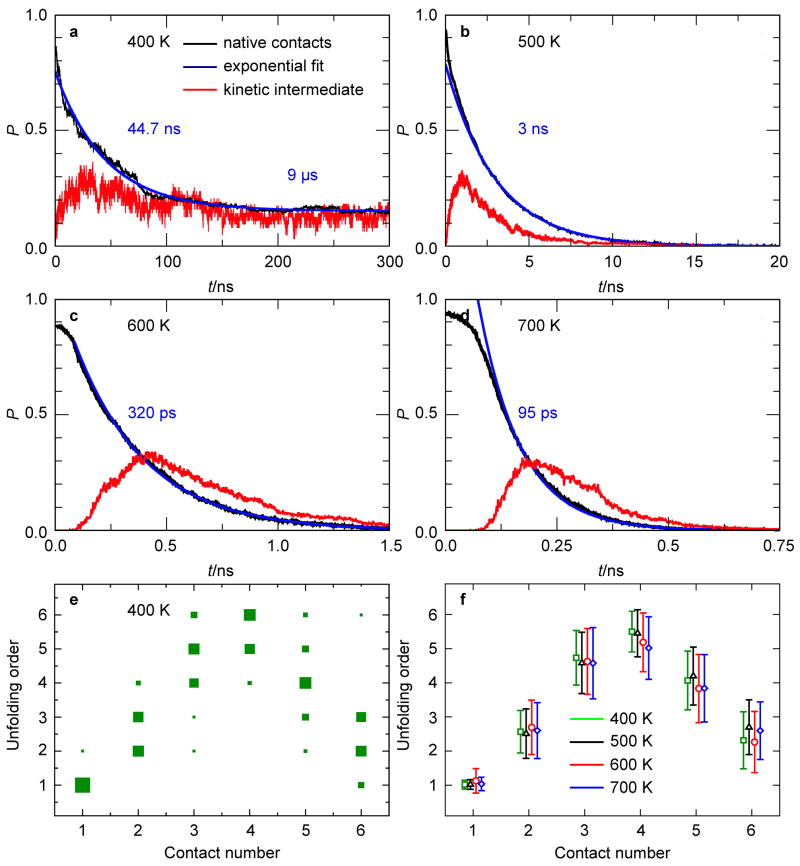
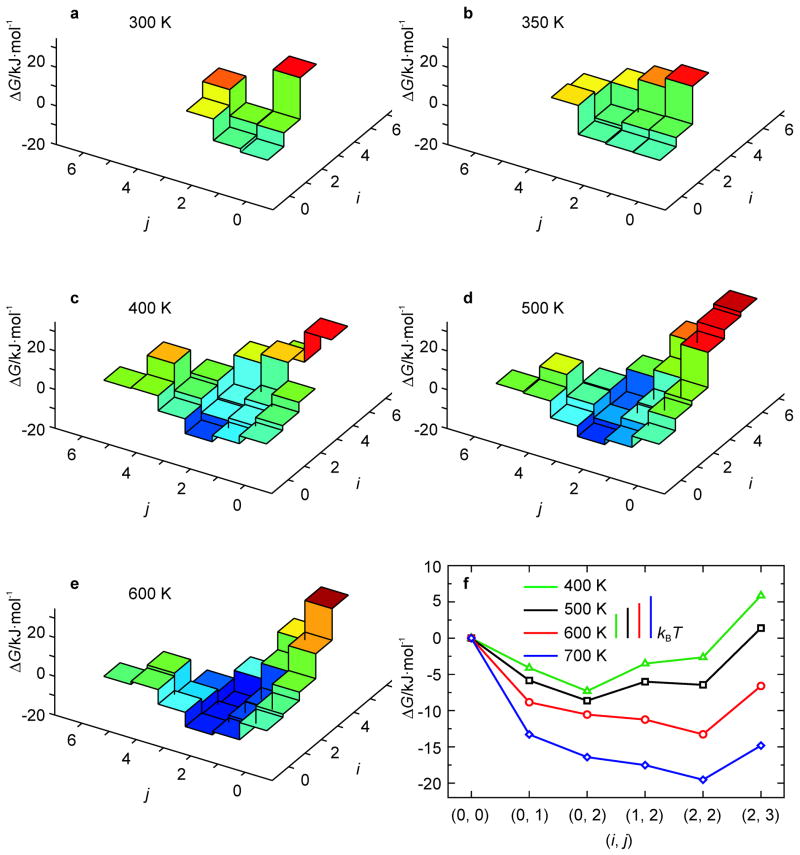
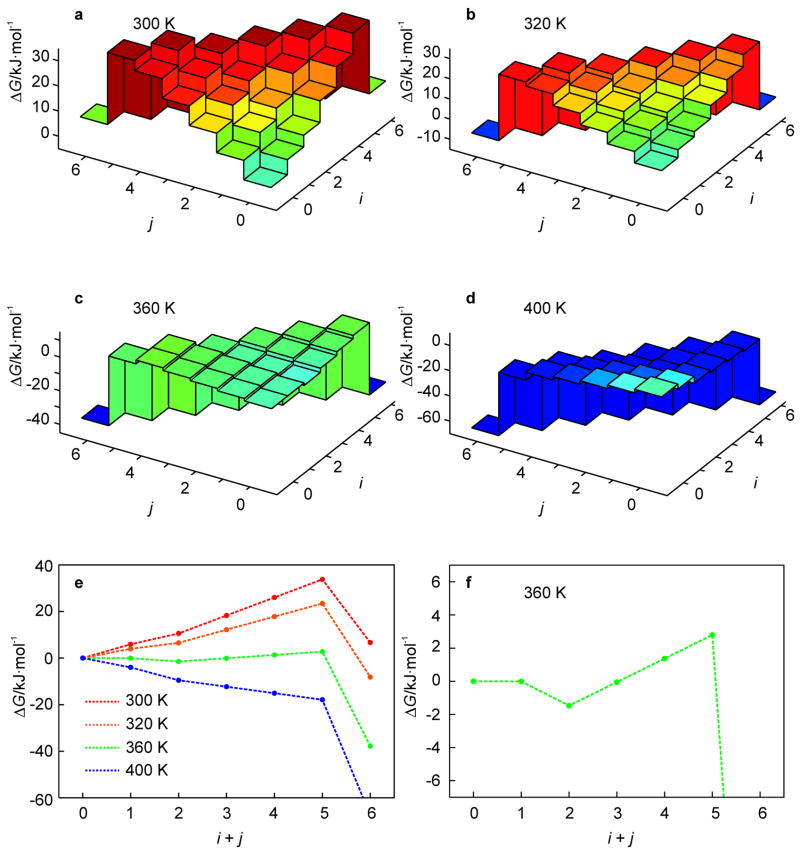
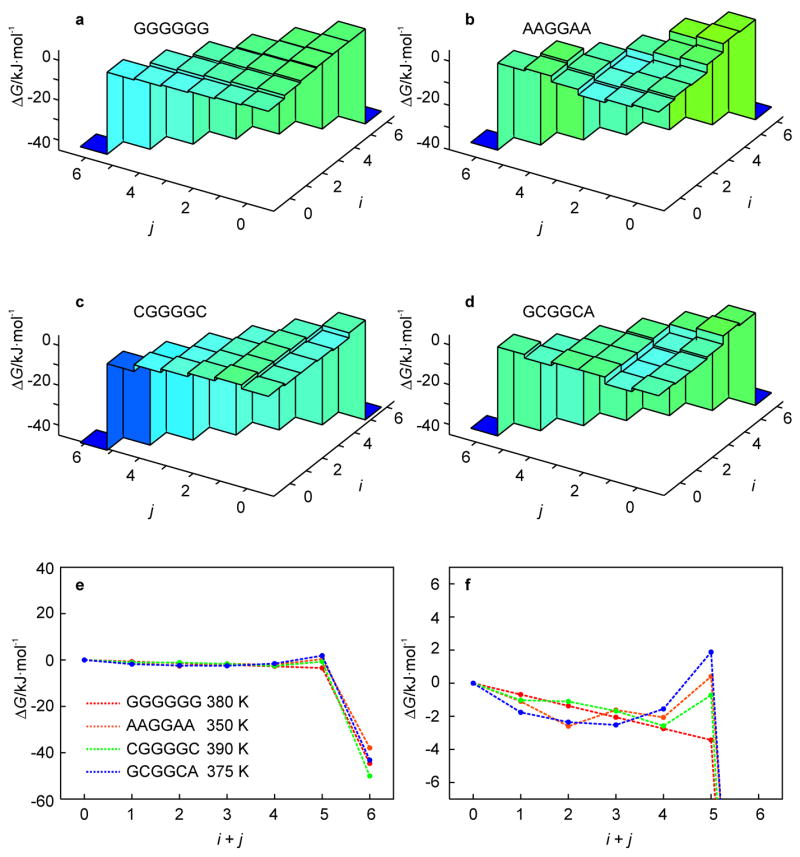
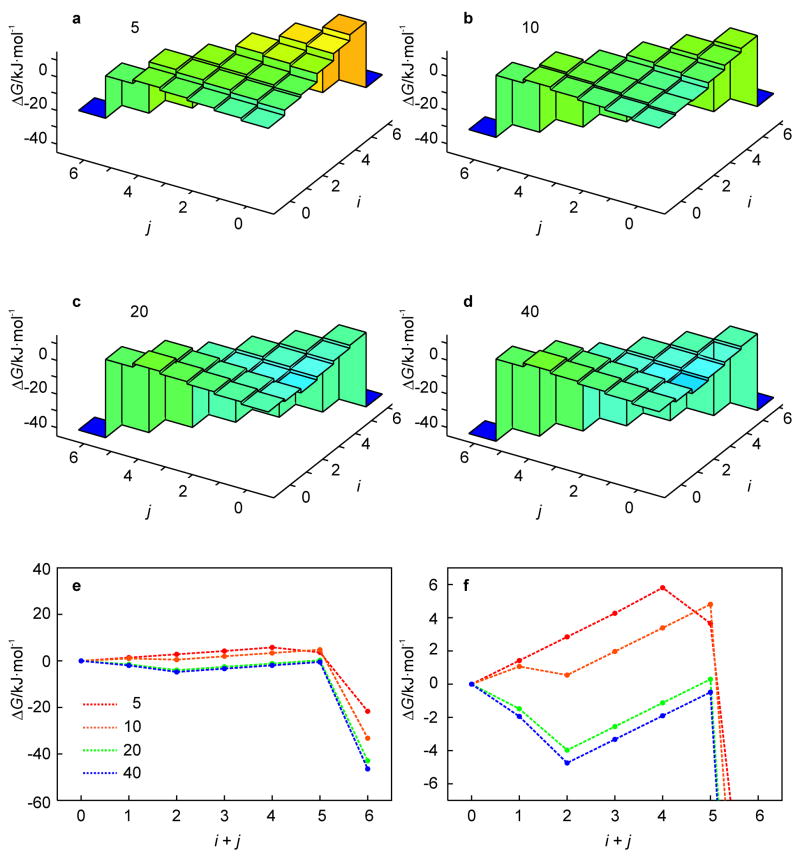
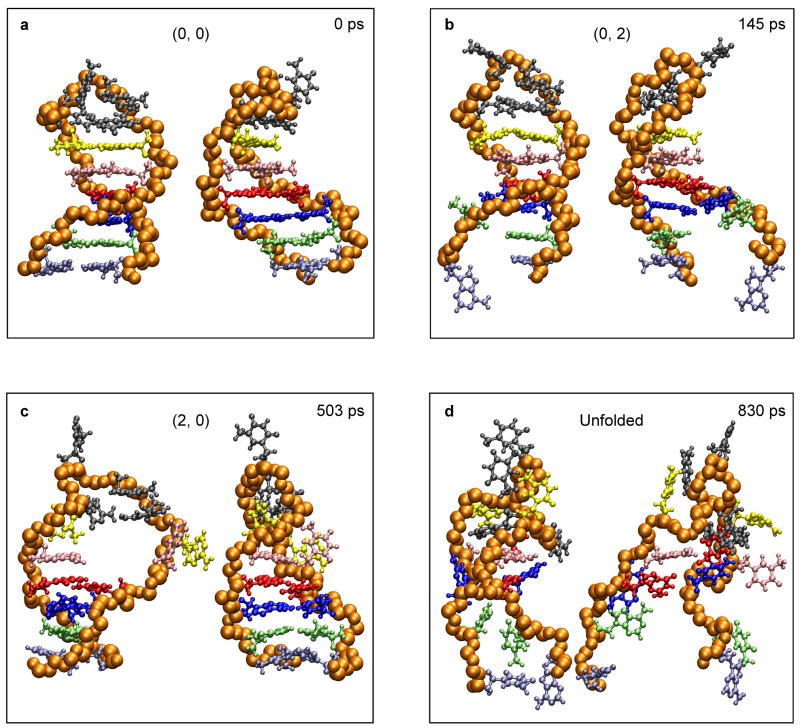
A 2D free-energy landscape model is presented to describe the (un)folding transition of DNA/RNA hairpins, together with molecular dynamics simulations and experimental findings. The dependence of the (un)folding transition on the stem sequence and the loop length is shown in the enthalpic and entropic contributions to the free energy. Intermediate structures are well defined by the two coordinates of the landscape during (un)zipping. Both the free-energy landscape model and the extensive molecular dynamics simulations totaling over 10 mus predict the existence of temperature-dependent kinetic intermediate states during hairpin (un)zipping and provide the theoretical description of recent ultrafast temperature-jump studies which indicate that hairpin (un)zipping is, in general, not a two-state process. The model allows for lucid prediction of the collapsed state(s) in simple 2D space and we term it the kinetic intermediate structure (KIS) model.
Figures
References
-
- Brion P, Westhof E. Annu Rev Biophys Biomol Struct. 1997;26:113–137. - PubMed
-
- Varani G. Annu Rev Biophys Biomol Struct. 1995;24:379–404. - PubMed
-
- Glucksmann-Kuis MA, Dai X, Markiewicz P, Rothman-Denes LB. Cell. 1996;84:147–154. - PubMed
-
- Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell JE. Molecular Cell Biology. 4. W. H. Freeman; New York: 2000.
-
- Uhlenbeck OC. Nature. 1990;346:613–614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources