

Role of the commissural inhibitory system in vestibular compensation in the rat
- PMID: 18635647
- PMCID: PMC2614028
- DOI: 10.1113/jphysiol.2008.155291
Role of the commissural inhibitory system in vestibular compensation in the rat
Abstract
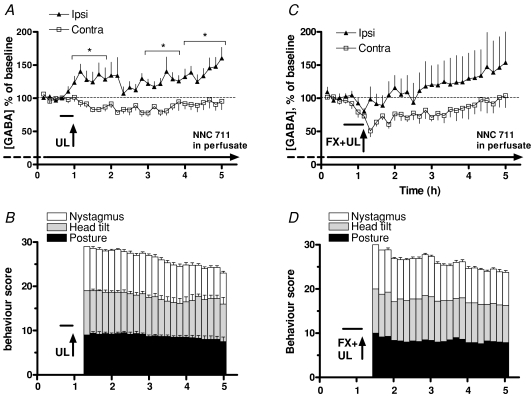
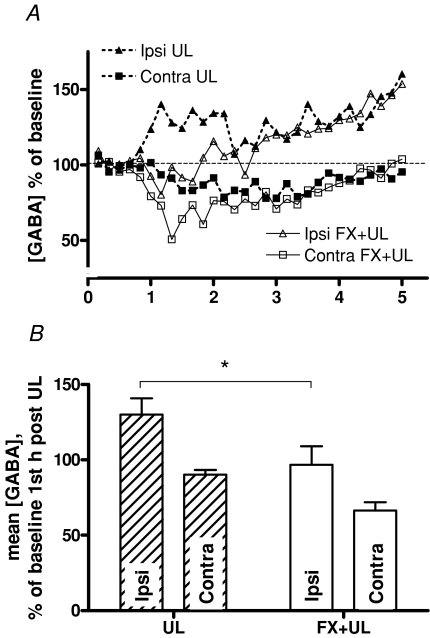
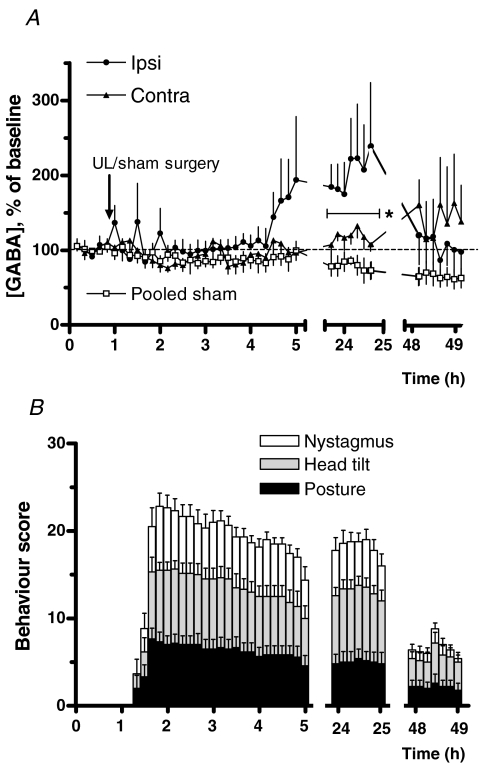
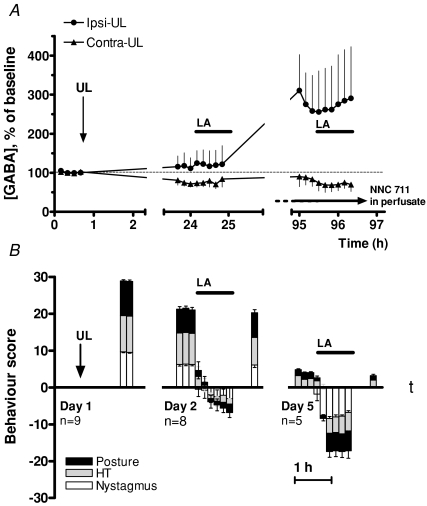
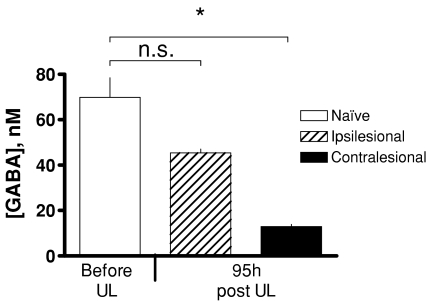
We investigated the role of the vestibular commissural inhibitory system in vestibular compensation (VC, the behavioural recovery that follows unilateral vestibular loss), using in vivo microdialysis to measure GABA levels in the bilateral medial vestibular nucleus (MVN) at various times after unilateral labyrinthectomy (UL). Immediately after UL, in close correlation with the appearance of the characteristic oculomotor and postural symptoms, there is a marked increase in GABA release in the ipsi-lesional MVN. This is not prevented by bilateral flocculectomy, indicating that it is due to hyperactivity of vestibular commissural inhibitory neurones. Over the following 96 h, as VC occurs and the behavioural symptoms ameliorate, the ipsi-lesional GABA levels return to near-normal. Contra-lesional GABA levels do not change significantly in the initial stages of VC, but decrease at late stages so that when static symptoms have abated there remains a significant difference between the MVNs of the two sides. We also investigated the role of the commissural inhibition in Bechterew's phenomenon, by reversibly inactivating the intact contra-lesional labyrinth in compensating animals through superfusion of local anaesthetic on the round window. Transient inactivation of the intact labyrinth elicited the lateralized behaviour described by Bechterew, but did not alter the GABA levels in either MVN, suggesting the involvement of distinct cellular mechanisms. These findings indicate that an imbalanced commissural inhibitory system is a root cause of the severe oculomotor and postural symptoms of unilateral vestibular loss, and that re-balancing of commissural inhibition occurs in parallel with the subsequent behavioural recovery during VC.
Figures
References
-
- Yu H, An Y, Jiang H, Jin Q, Jin Y. Changes of amino acid concentrations in the rat medial vestibular nucleus following unilateral labyrinthectomy. Sheng Li Xue Bao. 2007;59:71–78. - PubMed
-
- Barmack NH, Qian ZY, Kim HJ, Yoshimura J. Activity-dependent distribution of protein kinase C-delta within rat cerebellar Purkinje cells following unilateral labyrinthectomy. Exp Brain Res. 2001;141:6–20. - PubMed
-
- Beraneck M, Hachemaoui M, Idoux E, Ris L, Uno A, Godaux E, Vidal PP, Moore LE, Vibert N. Long-term plasticity of ipsilesional medial vestibular nucleus neurons after unilateral labyrinthectomy. J Neurophysiol. 2003;90:184–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources