

doi: 10.1126/science.1157632.
Evolutionary origins for social vocalization in a vertebrate hindbrain-spinal compartment
Affiliations
- PMID: 18635807
- PMCID: PMC2582147
- DOI: 10.1126/science.1157632
Item in Clipboard
Evolutionary origins for social vocalization in a vertebrate hindbrain-spinal compartment
Science.
.
Abstract
The macroevolutionary events leading to neural innovations for social communication, such as vocalization, are essentially unexplored. Many fish vocalize during female courtship and territorial defense, as do amphibians, birds, and mammals. Here, we map the neural circuitry for vocalization in larval fish and show that the vocal network develops in a segment-like region across the most caudal hindbrain and rostral spinal cord. Taxonomic analysis demonstrates a highly conserved pattern between fish and all major lineages of vocal tetrapods. We propose that the vocal basis for acoustic communication among vertebrates evolved from an ancestrally shared developmental compartment already present in the early fishes.
Figures
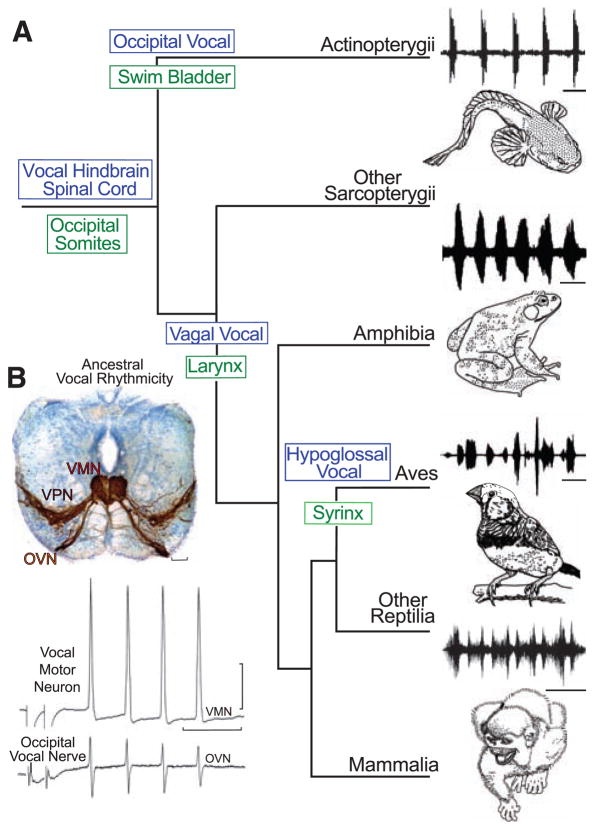
Evolution of vocal behaviors. (A) Cladogram of living bony vertebrates (3), with oscillogram of a vocalization from a representative species, shows nodal (ancestral) states for vocal characters (10). Vocalizations (top to bottom): midshipman fish agonistic “grunts,” bullfrog advertisement call, estrildid finch song, and squirrel monkey cackle. Scale bars (top to bottom) are 500 ms, 1.0 s, 250 ms, and 200 ms. Vocal mechanisms are unknown for lobe-finned fish (other Sarcopterygii), although well-known for nonavian (other) Reptilia (30). (B) Vocal pacemaker circuit. Among batrachoidid fish (midshipman and toadfish), there is a direct translation between the temporal properties of the vocal circuitry and natural calls (4, 5). (Top) Transverse section at the caudal hindbrain-spinal cord transition of a toadfish shows transneuronal, neurobiotin-labeling of midline vocal motor neurons (VMNs), adjacent pacemaker neurons (VPN), and motor axons exiting via nerve root that gives rise to the occipital vocal nerve (OVN); vocal neurons have extensive lateral processes (5, 6). Scale bar is 100 μm. (Bottom) The rhythmic, oscillatory-like activity of a vocal motor neuron [(top trace) average of four DC-coupled intracellular records] is aligned with occipital nerve activity [(bottom trace) average of four intracranial records] to indicate relative timing [see (5) for same temporal pattern of pacemaker neurons]. Response was evoked by midbrain electrical microstimulation in midshipman fish [paired stimulus artifact far left; modified from (5) with permission]. Horizontal scale bar is 20 ms; vertical scale bar is 20 mv and 1 mv, respectively, for intracellular and nerve records.
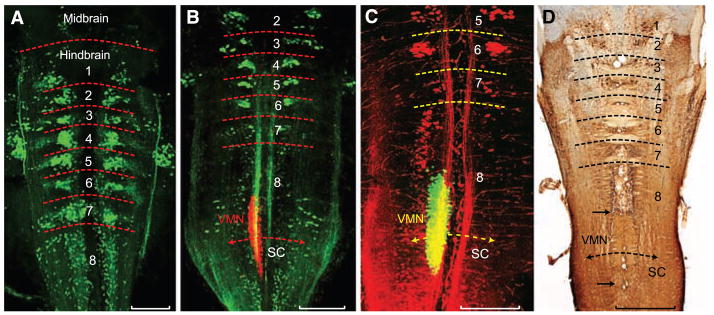
Segmental organization of hindbrain reticulospinal and vocal neurons. (A to C) Confocal projections in horizontal plane of fluorescently labeled neurons in midshipman larvae [n = 21; 8 to 14 days post fertilization (dpf), 7- to 10-mm standard length]. Horizontal hatching approximates midbrain, hindbrain rhombomere (1 to 8), and spinal cord (SC) boundaries. Scale bars are 0.2 mm. (A) Clusters of reticulospinal neurons labeled with Alexa biocytin 488 (green) at a stage before vocal muscle reaches swim bladder (~8 dpf). (B) Simultaneous visualization of reticulospinal neurons and vocal motor neurons (VMNs) labeled with, respectively, Alexa 488 dextran-amine (green) and Alexa 546 dextran-amine (red) at a stage when vocal muscles attach to the swim bladder (~14 dpf). (C) Same stage as (B) highlights rostral subdivision of rh8 reticulospinal column (Alexa 546 dextran-amine, red) and VMN (Alexa 488 dextran-amine, green). Yellow in (B) and (C) is composite overlap (red and green), but no double label. (D) Horizontal section showing biocytin-filled, reticulospinal clusters in postlarval midshipman (80 mm, ~200 dpf). Paired black arrows denote VMN’s rostral-caudal extent. Scale bar is 0.5 mm.
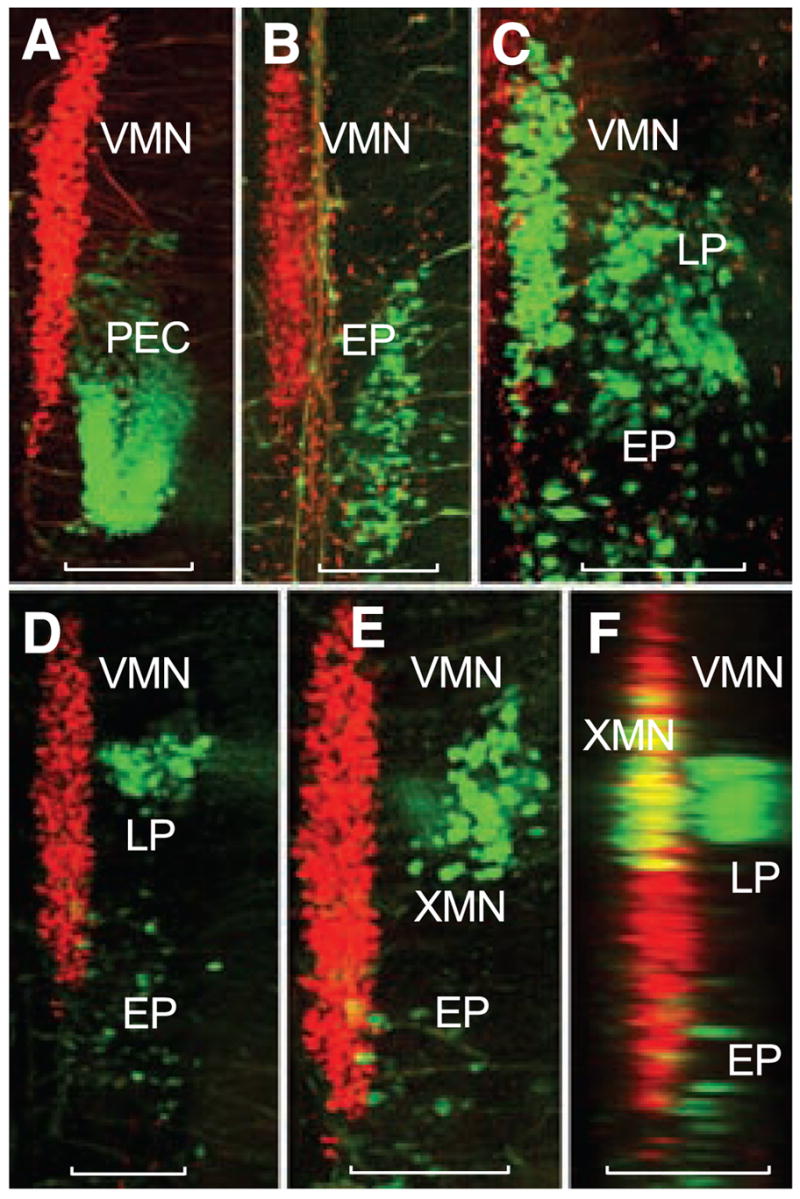
Segmental organization of vocal and non-vocal motor neurons. Simultaneous labeling of vocal and either nonvocal muscles or nerve roots with different wavelength fluorescent dextran-amines provides visualization of vocal motor neurons (VMNs) and nonvocal motor neurons in midshipman larvae (~14 dpf, n = 28). Confocal projections in horizontal (A to E) and sagittal (F) planes. Nonvocal motor neurons are designated by target muscle: EP, epaxialis; LP, levator pectoralis; PEC, pectoral fin. XMN indicates vagal motor neurons innervating wall of gastrointestinal tract. Neurons appearing red and green were labeled with, respectively, fluorescent dextran-amines 546 and 488. Scale bars are 0.1 mm. (A and B) PEC and EP neurons lateral to caudal vocal motor neurons. (C) LP and EP lateral to caudal vocal motor neurons after intracranial label of ipsilateral occipital roots; partial view of contralateral vocal motor neurons filled (far left) by labeling contralateral occipital root. (D) LP neurons lateral to vocal motor neurons with sparsely distributed EP neurons labeled during the exposure. (E) Higher magnification view just dorsal to (D) showing XMN lateral, rather than directly dorsal, to vocal motor neurons because of tissue flat mounting (see fig. S1). (F) Sagittal reconstruction of (D) and (E) showing XMN and LP relative to more medial vocal motor neurons. Yellow is composite overlap (red and green), but no double label.
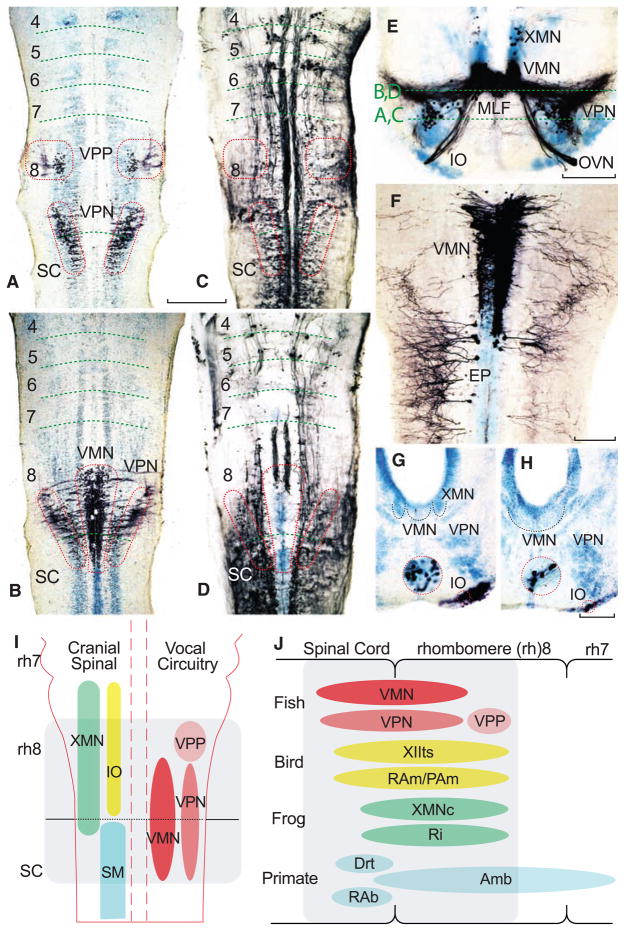
Segmental organization of vocal pacemaker–motor neuron network along with summary diagrams of larval fish and vocal vertebrates. (A to H) Alignment of either neurobiotin- or dextran-biotin–filled (which appear black) vocal and nonvocal neurons in toadfish (A to F) and midshipman (G and H) larvae (n = 25; 1.8 to 2.0 cm, ~30 to 40 dpf). Cresyl violet counterstain illustrates other neuronal groups. Paired horizontal sections at two dorsal-ventral levels, indicated in (E), compare neurobiotin-filled vocal prepacemaker (VPP), pacemaker (VPN), and motor (VMN) neurons (A and B) relative to neurobiotion-filled reticulospinal neurons (C and D). Red hatching in (C) and (D) approximates location of the VPP, VPN, and VMN in (A) and (B). Horizontal hatching in (A to D) estimates rhombomere (4 to 8) and spinal cord (SC) boundaries. Scale bar is 0.2 mm for (A to D). (E) Transverse section through rostral, neurobiotin-filled vocal pacemaker–motor neurons and exiting occipital vocal tract and nerve root (OVN), including a few labeled vagal motor neurons (XMN) that innervate swim bladder smooth muscle (15). (IO, inferior olive; MLF, medial longitudinal fasciculus.) Scale bar is 0.1 mm. (F) Horizontal section of VMN (right side) and trunk (epaxialis, EP; left side) motor neurons labeled with 10-kD biotin-dextran-amine. VMN exhibits dendritic spread ipsilaterally and into contralateral VMN. Scale bar is 0.1 mm. (G and H) Transverse sections showing neurobiotin-labeled inferior olive (IO) coextensive with rostral (G) and middle (H) VMN levels (cresyl violet–stained VMN, outlined with black dotted line, is adjacent to XMN, also outlined, and/or VPN). Red-ringed inserts are higher magnification views of IO. Scale bars are 0.1 and 0.03 mm. (I) Schematic in horizontal plane showing relative positions of vocal and nonvocal neurons comprising a rhombomere (rh) 8 to spinal (SC) compartment. (J) Comparative summary in sagittal plane showing relative positions of vocal neurons in the rh8–spinal compartment. The developmental map for premotor-motor circuitry in fish (this report), as well as for motor neurons in birds (13) and frogs (17), is based on representatives of similar age. Positioning of premotor neurons that participate in vocal patterning in birds (20, 21) and frogs (22), and motor and premotor neurons in mammals (19, 23, 24), including primates (25), is based on adult phenotypes [but see (8) for developmental studies of caudal hindbrain in mammals]. Most laryngeal motor neurons that shape the temporal envelope of mammalian calls originate from caudal nucleus ambiguus (Amb) (23). Abbreviations: Drt, dorsal reticular nucleus; PAm, nucleus parambigualis; Ri, inferior reticular formation; RAb, nucleus retroambiguus; RAm, nucleus retroambigualis; VPP-VPN-VMN, vocal prepacemaker–pacemaker–motor neurons; XMNc, caudal XMN; XIIts, tracheosyringeal division of hypoglossal motor nucleus.
Comment in
-
Neuroscience. Vertebrate vocalizations.Science. 2008 Jul 18;321(5887):347-8. doi: 10.1126/science.1161775. Science. 2008. PMID: 18635781 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
