

The tumour suppressor CYLD is a negative regulator of RIG-I-mediated antiviral response
- PMID: 18636086
- PMCID: PMC2529351
- DOI: 10.1038/embor.2008.136
The tumour suppressor CYLD is a negative regulator of RIG-I-mediated antiviral response
Abstract
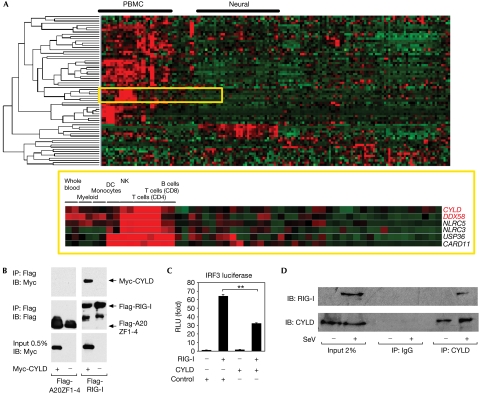
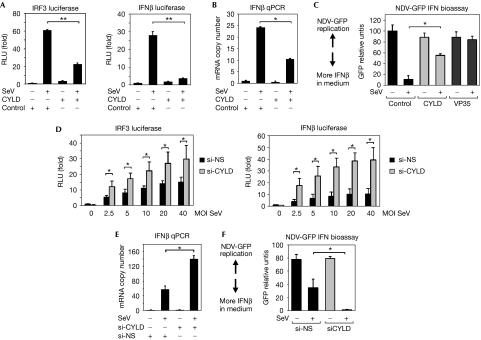
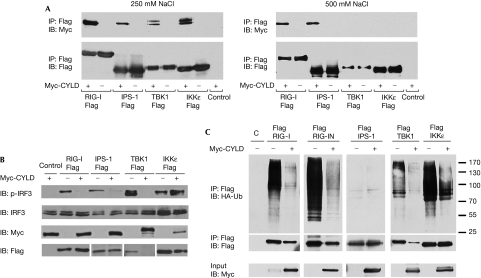
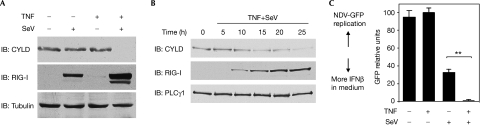
On detecting viral RNAs, the RNA helicase retinoic acid-inducible gene I (RIG-I) activates the interferon regulatory factor 3 (IRF3) signalling pathway to induce type I interferon (IFN) gene transcription. How this antiviral signalling pathway might be negatively regulated is poorly understood. Microarray and bioinformatic analysis indicated that the expression of RIG-I and that of the tumour suppressor CYLD (cylindromatosis), a deubiquitinating enzyme that removes Lys 63-linked polyubiquitin chains, are closely correlated, suggesting a functional association between the two molecules. Ectopic expression of CYLD inhibits the IRF3 signalling pathway and IFN production triggered by RIG-I; conversely, CYLD knockdown enhances the response. CYLD removes polyubiquitin chains from RIG-I as well as from TANK binding kinase 1 (TBK1), the kinase that phosphorylates IRF3, coincident with an inhibition of the IRF3 signalling pathway. Furthermore, CYLD protein level is reduced in the presence of tumour necrosis factor and viral infection, concomitant with enhanced IFN production. These findings show that CYLD is a negative regulator of RIG-I-mediated innate antiviral response.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Bignell GR et al. (2000) Identification of the familial cylindromatosis tumour-suppressor gene. Nat Genet 25: 160–165 - PubMed
-
- Brummelkamp TR, Nijman SM, Dirac AM, Bernards R (2003) Loss of the cylindromatosis tumour suppressor inhibits apoptosis by activating NF-κB. Nature 424: 797–801 - PubMed
-
- Fitzgerald KA, McWhirter SM, Faia KL, Rowe DC, Latz E, Golenbock DT, Coyle AJ, Liao SM, Maniatis T (2003) IKKɛ and TBK1 are essential components of the IRF3 signaling pathway. Nat Immunol 4: 491–496 - PubMed
-
- Gack MU et al. (2007) TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature 446: 916–920 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30 DK040561/DK/NIDDK NIH HHS/United States
- AI062623/AI/NIAID NIH HHS/United States
- R21 AI057997/AI/NIAID NIH HHS/United States
- AI073919/AI/NIAID NIH HHS/United States
- T32 AI07647/AI/NIAID NIH HHS/United States
- AI057997/AI/NIAID NIH HHS/United States
- AI057158/AI/NIAID NIH HHS/United States
- R01 AI062773/AI/NIAID NIH HHS/United States
- R56 AI052417/AI/NIAID NIH HHS/United States
- U19 AI062623/AI/NIAID NIH HHS/United States
- AI065058/AI/NIAID NIH HHS/United States
- AI062773/AI/NIAID NIH HHS/United States
- T32 AI007647/AI/NIAID NIH HHS/United States
- R01 AI041111/AI/NIAID NIH HHS/United States
- AI041111/AI/NIAID NIH HHS/United States
- R01 AI073919/AI/NIAID NIH HHS/United States
- R01 AI052417/AI/NIAID NIH HHS/United States
- F32 AI065058/AI/NIAID NIH HHS/United States
- AI052417/AI/NIAID NIH HHS/United States
- AI059536/AI/NIAID NIH HHS/United States
- R01 AI059536/AI/NIAID NIH HHS/United States
- U54 AI057158/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
