

Role of the hemopexin domain of matrix metalloproteinases in cell migration
- PMID: 18636552
- PMCID: PMC2574584
- DOI: 10.1002/jcp.21535
Role of the hemopexin domain of matrix metalloproteinases in cell migration
Abstract
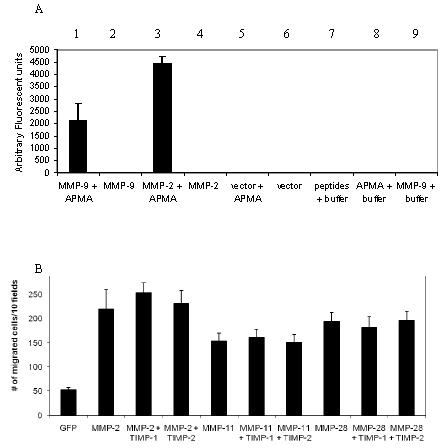
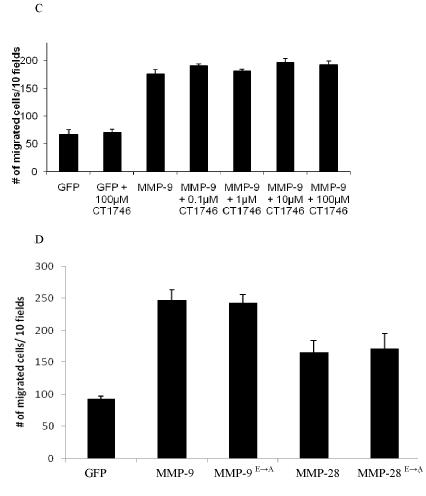
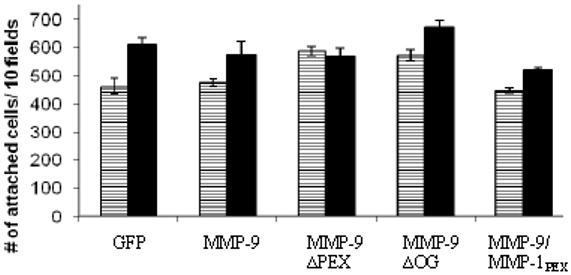
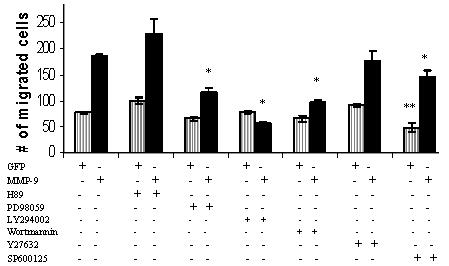
The biological functions of matrix metalloproteinases (MMPs) extend beyond extracellular matrix degradation. Non-proteolytic activities of MMPs are just beginning to be understood. Herein, we evaluated the role of proMMPs in cell migration. Employing a Transwell chamber migration assay, we demonstrated that transfection of COS-1 cells with various proMMP cDNAs resulted in enhancement of cell migration. Latent MMP-2 and MMP-9 enhanced cell migration to a greater extent than latent MMP-1, -3, -11 and -28. To examine if proteolytic activity is required for MMP-enhanced cell migration, three experimental approaches, including fluorogenic substrate degradation assay, transfection of cells with catalytically inactive mutant MMP cDNAs, and addition of hydroxamic acid-derived MMP inhibitors, were employed. We demonstrated that the proteolytic activities of MMPs are not required for MMP-induced cell migration. To explore the mechanism underlying MMP-enhanced cell migration, structure-function relationship of MMP-9 on cell migration was evaluated. By using a domain swapping approach, we demonstrated that the hemopexin domain of proMMP-9 plays an important role in cell migration when examined by a transwell chamber assay and by a phagokinetic migration assay. TIMP-1, which interacts with the hemopexin domain of proMMP-9, inhibited cell migration, whereas TIMP-2 had no effect. Employing small molecular inhibitors, MAPK and PI3K pathways were found to be involved in MMP-9-mediated cell migration. In conclusion, we demonstrated that MMPs utilize a non-proteolytic mechanism to enhance epithelial cell migration. We propose that hemopexin homodimer formation is required for the full cell migratory function of proMMP-9.
Figures







References
-
- Abecassis I, Olofsson B, Schmid M, Zalcman G, Karniguian A. RhoA induces MMP-9 expression at CD44 lamellipodial focal complexes and promotes HMEC-1 cell invasion. Exp. Cell. Res. 2003;291:363–376. - PubMed
-
- Albrect-Buehler G. The phagokinetic tracks of 3T3 cells. Cell. 1997;11:395–404. - PubMed
-
- Bannikov GA, Karelina TV, Collier IE, Marmer BL, Goldberg GI. Substrate binding of gelatinase B induces its enzymatic activity in the presence of intact propeptide. J. Biol. Chem. 2002;277:16022–16027. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
